 Гаплогруппа
Гаплогруппа
(в
популяционной генетике человека, науке, изучающей генетическую историю
человечества) — большая группа схожих гаплотипов, которые являются рядом
аллелей на нерекомбинируемых участках Y-хромосомы. Галпогруппы делятся
на Y-хромосомные (Y-ДНК) и митохондриальные (мт-ДНК). Y-ДНК является
прямой отцовской линией, т.е сын, отец, дед и т.д., а мт-ДНК прямой
материнской линией, то есть дочь, мать, бабушка, прабабушка и так далее.
Термин «гаплогруппа» широко применяется в генетической ДНК генеалогии.
Гаплогруппа R1a
Гаплогруппа R1a1 насчитывает около 300 миллионов мужчин. Первый общий предок современных носителей R1a1 жил около 300 поколений назад.
Распространение гаплогруппы R1a :
В процентах указана доля R1a от общей численности этноса
- Русские 48%
- Поляки 56%
- Украинцы 54%
- Белорусы 51%
- Чехи 34%
- Киргизы 63%
- Шорцы 56%
- Алтайцы 54%
- Чуваши 31,5%
- Таджики 53%
- Панджабцы 54% (Пакистан-Индия)
- Индия в целом 30%, верхние касты 43%
Экскурс в Древнюю историю гаплогруппы R1a
Гаплогруппа R1a1 возникла
около 15 000 лет назад в Азии и в дальнейшем распалась на несколько
субкладов, или как их еще называют, дочерних гаплогрупп. Мы рассмотрим
основные из них - это Z283 и Z93. R1a1-Z93 - это азиатский маркер,
характерный для тюрков, евреев, индийцев. С участием гаплогруппы
R1a1-Z93 изобрели колесо в степи, сконструировали первые повозки и
приручили лошадь. Это были культуры Андроновского круга. Гаплогруппа
быстро освоила всю полосу евразийских степей от Каспия до Забайкалья,
распавшись на множество различных племен с разными этнокультурными
особенностями.
R1a1-Z283 - это европейский маркер и характерен по большей части для
славян, но не только, свои отдельные субклады имеются и у скандинавов,
англичан. В целом на сегодняшний день древняя гаплогруппа R1a1 наиболее характерна для славянских, тюркских и индийских этносов.

Раскопки "Страны городов" на Южном Урале подтвердили, что уже примерно
4000 лет назад в укрепленном поселении Аркаим имелись помещения личного и
общественного пользования, жилые и мастерские. В некоторых помещениях
обнаружены не только гончарные мастерские, но и металлургическое
производство.
В ходе раскопок было вскрыто около 8000 кв. м площади поселения
(примерно половина), вторая часть была исследована с помощью
археомагнитных методов. Тем самым была полностью установлена планировка
памятника. Здесь же был впервые в Зауралье применён метод реконструкции,
и Л.Л. Гуревичем сделаны рисунки возможного вида поселения. R1a1-Z93
вероятно была одной из основных гаплогрупп в Аркаиме.
Многие племена, вступая в контакт с этими общинами постепенно переходили
на их язык. В настоящее время почти вся Европа говорит на
индоевропейских языках, в то время как гаплогруппа R1b более специфична западной Европе, а R1a - восточной Европе. В странах находящихся ближе к центральной Европе имеются обе эти гаплогруппы. Так гаплогруппа R1a занимает
около 30% в населении Норвегии, и небольшой процент (не более 15%) в
Восточной Германии - остатки прямых Y-линий некогда ассимилированных
немцами полабских славян.

Во втором тысячелетии до нашей эры предположительно из-за климатических
изменений часть R1a1 (субклад Z93 и др. гаплогруппы Средней Азии) начали
мигрировать на юг и восток за пределы степи, часть (субклад L657) пошла
в сторону Индии и, влившись в местные племена, приняла участие в
создании кастового общества. Те далекие события описаны в древнейшем
литературном источнике человечества –"Ригведе".
Другая часть стала продвигаться в ближневосточном направлении. На территории современной Турции предположительно они основали Хеттское государство,
успешно конкурировавшее и с древним Египтом. Хетты строили города, но
не смогли прославиться сооружением огромных пирамид, так как в отличие
от Египта Хеттское общество было обществом свободных людей, и им
были чужды идеи использования подневольного труда. Хеттское государство
исчезло внезапно, его смела мощная волна варварских племен известных
как "народы моря". В середине прошлого века археологи нашли богатейшую
библиотеку глиняных табличек с хеттскими текстами, язык оказался
относящимся к индо-европейской группе языков. Так мы обрели детальные
знания о первом государстве, часть мужских линий которого
предположительно состояла из гаплогруппы R1a1-Z93.
Славянские субклады гаплогруппы R1a1-Z283 образуют свой кластер гаплотипов, который совершенно не родственен ни западно-европейским субкладам гаплогруппы R1a, ни индо-иранским и разделение европейских носителей R1a1-Z283 с азиатскими R1a1-Z93 произошло примерно 6 500 лет назад.

В октябре 539 года (до нашей эры) иранское племя Персов захватило Вавилон, вождь персов Кир решил
не уходить, а серьезно обосноваться в захваченном городе. Впоследствии
Киру удалось значительно расширить свои владения, так возникла великая
Персидская Империя, просуществовавшая дольше всех империй в мире - 1190
лет! В 651 году нашей эры ослабленная междоусобицей Персия пала под
натиском арабов, и это привело к изменению гаплогруппного состава
населения. Сейчас на обширных территориях, занимаемых некогда иранским
государством, практически не осталосьгаплогруппы R1a, по иронии судьбы Иран, остался без гаплогруппы, внесшей большой вклад на этапе становления этого государства.
Три мировые религии связаны с индо-ариями - Индуизм, Буддизм и Зороастризм.
Зороастр был персом, а
Будда происходил из племени индусов
Саков. Возможно они были носителями гаплогруппы R1a1.
Большинство народов состоит из множества гаплогрупп, и не имеется рода,
который бы доминировал над остальными. Связь между гаплогруппой и
внешностью человека также отсутствует и как видно многие представители гаплогруппы R1a1 относятся даже к разным расам. Многим R1a1-Z93
характерны монголоидные черты (киргизы, алтайцы, хотоны и т.д.), в то
время как носители R1a1-Z283 имеют в большинстве европейский вид
(поляки, русские, белорусы и т.д.). Большое количество финских племен
имеют высокие проценты гаплогруппы R1a1, часть из которых были ассимилированы с приходом славянских колонистов 9 века.
Достижения, к которым возможно имеют отношение R1a1:
Колесо, повозки, приручение лошади, металлургия, брюки, сапоги, платья,
первый в мире мощеный "автобан" протяженностью более 1000 км со
станциями "заправки" - замены лошадей, и много чего другого.
В небольшой статье трудно рассказать всю историю первых индо-европейцев,
можно лишь некоторыми историческими фрагментами пробудить интерес к
истории древних предков славян. Наберите в поисковике слова индо-арии, тюрки, славяне,скифы, сарматы, Персия, и Вы окунетесь в увлекательное путешествие по славной истории индо-европейских и славянских народов.
Древо гаплогруппы.
До 2007 года никто не осуществляли детальных реконструкций родов, никому
в голову не пришла эта идея, да и решить такую грандиозную задачу не
было возможности. Многие популяционные генетики работали с небольшими
выборками коротких 6-маркерных гаплотипов, позволяющих получать общие
генографические представления о распределении гаплогрупп.
 В
2009 году профессиональный популяционный генетик задался целью
построить детальное генеалогическое древо этой гаплогруппы. Столкнувшись
с рядом проблем, например, расчет обычными методами крупных выборок
сверх длинных гаплотипов был невозможен из-за астрономического
количества операций, ни один компьютер был не в состоянии перебрать
нужное количество комбинаций, но благодаря находчивости и желанию
построить древо своей гаплогруппы проблемы эти удалось преодолеть.
В
2009 году профессиональный популяционный генетик задался целью
построить детальное генеалогическое древо этой гаплогруппы. Столкнувшись
с рядом проблем, например, расчет обычными методами крупных выборок
сверх длинных гаплотипов был невозможен из-за астрономического
количества операций, ни один компьютер был не в состоянии перебрать
нужное количество комбинаций, но благодаря находчивости и желанию
построить древо своей гаплогруппы проблемы эти удалось преодолеть.
После R1a1 многие гаплогруппы приступили к созданию своих древ.
Сами гаплогруппы не несут генетической информации, т.к. генетическая
информация находиться в аутосомах- первых 22 пар хромосом. Посмотреть
распространение генетических компонент в Европе можно здесь. Гаплогруппы являются всего лишь метками давно минувших дней, на заре формирования современных народов.
Гаплогруппа R1b
Гаплогруппа
R1b – параллельный субклад для гаплогруппы R1a. Родоначальник
гаплогруппы R1b родился около 16 000 лет назад в центральной Азии от
родительского рода R1. Примерно 10 000 лет назад гаплогруппа R1b
распалась на несколько субкладов, которые начали расходиться в разных
направлениях. Восточную ветвь - субклад R1b-M73 некоторые ученые
связывают с древними тохарами, которые приняли участие в этногенезе
такого народа, как современные уйгуры.

Продвижение гаплогруппы R1b на
запад в Европу вероятно происходило в несколько этапов. Часть может
быть связана с неолитическими миграциями из Малой Азии и Закавказья, а
часть с миграциями постнеолитическими и распространением археологической
культуры Колоколовидных Кубков. Существует версия и о миграции по
Североафриканскому побережью к Гибралтарскому проливу, с дальнейшей
переправкой на Пиренеи в виде археологической культуры Колоколовидных
кубков - но в данной гипотезе слишком много натяжек. В любом случае
большинство европейских представителей гаплогруппы R1b имеют снип P312,
который определенно произошел уже в Европе.
После того, как египетские ученые провели анализ мумии Тутанхамона, было обнаружено, что фараон оказался представителем гаплогруппы R1b.
Сейчас основная масса представителейгаплогруппы R1b1a2 проживает в Западной Европе, где гаплогруппа R1b1a2 является
основной гаплогруппой. В России только башкирский народ имеет большой
процент этой гаплогруппы. В русском народе гаплогруппа R1b составляет не
более 5%. В Петровскую и Екатерининскую эпоху проводилась
государственная политика массового привлечения иностранных специалистов
из Германии и остальной Европы, многие русские R1b являются их
потомками. Также какая-то часть могла войти в русский этнос с Востока -
это прежде всего субклад R1b-M73. Некоторые R1b-L23 могут быть
мигрантами с Кавказа, куда они попали из Закавказья и Передней Азии.
Европа
Современная концентрация гаплогруппы R1b максимальна
на территориях миграционных путей кельтов и германцев: в южной Англии
около 70%, в северной и западной Англии, Испании, Франции, Уэльсе,
Шотландии, Ирландии — до 90 % и более. А так же, например, у басков — 88,1 %, испанцев — 70 %, итальянцев — 40 %, бельгийцев — 63 %, немцев — 39 %, норвежцев — 25,9 % и других.
В Восточной Европе гаплогруппа R1b встречается намного реже. У чехов и словаков — 35,6 %, латышей — 10 %, венгров — 12,1 %, эстонцев — 6 %, поляков — 10,2 %-16,4 %, литовцев — 5 %, белорусов — 4,2 %, русских — от 1,3 % до 14,1 %,украинцев — от 2 % до 11,1 %.
На Балканах — у греков — от 13,5 % до 22,8 %, словенцев — 21 %, албанцев — 17,6 %, болгар — 17 %, хорватов — 15,7 %,румын — 13 %, сербов — 10,6 %, герцеговинцев — 3,6 %, боснийцев — 1,4 %.
Азия
На Южном Урале значительно распространена у башкир — около 43 %.
На Кавказе обнаружена у осетин Дигора — 23 % и армян — 28,4 %.
В Турции достигает 16,3 %, Ираке — 11,3 % и в других странах Западной Азии.
В Центральной Азии обнаружена, в частности, у туркменов — 36,7 %, узбеков — 9,8 %, татар — 8,7 %, казахов — 5,6 %,уйгуров — от 8,2 % до 19,4 %
Африка
Гаплогруппа N

Гаплогруппа N возникла около 15 000 лет назад на юго-западе современного
Китая (провинция Юннань), и около 10 000 лет назад начала свой путь
движения в сторону Байкала, Саян и Алтая. На Байкале часть
представителей гаплогруппы N осталась, часть направилась на северо-запад
в сторону Балтики, а другая часть в сторону Дальнего Востока. Пройдя
через всю Сибирь, Урал и Русскую равнину, племена достигли побережья
Балтийского моря около 4 000 лет назад предположительно в рамках
археологической культуры Ямочно-Гребенчатой Керамики.

На всем пути продвижения на базе рода формировались разные этносы, по
большей части финно-угорские, тюркские, славянские и балтские (снип
L1025). На сегодняшний день максимальные частоты N1c1 обнаружены у
финнов и якутов, у которых гаплогруппа N1с1 достигает 80%, а
также балты (латыши и литовцы) - у них N1c1 достигает 47%. Современные
народы у которых преобладает общая гаплогруппа зачастую совершенно
различны антропологически, т.к. имеют разные субклады, разделенные около
5-7 тыс. лет назад, это объясняется просто: осваивать новые земли
отправлялись здоровые, молодые мужчины, а старики, женщины и дети
оставались на месте. Молодые войны берут жен из местных племен и таким
образом, каждое новое поколение, произошедшее от европейских женщин,
становилось все более и более европейским по внешнему виду.
В русском этносе гаплогруппа N занимает почетное второе место по распространенности. Наличие гаплогруппы N1с1 в
генофонде русских объясняется с одной стороны наличием субклада N1c1
L550 у славян (включая западных) а также ассимиляцией некоторых
финно-угорских народов славянскими колонистами 9 века; наличие
гаплогруппы N1c1 у поволжских народов тоже достигает высоких процентов.
Такие народы как мурома, полностью растворились в русских, но их мужские
линии продолжились в русском и других этносах до настоящего времени.

Гаплогруппа N1 прослеживается также в южном Китае, Бирме, Тайланде, Камбодже, Японии, Тайване и Корее.
Результаты дДНК подтвердили наличие N1 в останках культуры
Яншао.
Возникновение тюркских языков многие исследователи также связывают с
гаплогруппой N1, возможно N1b. Реконструкция времени разделения
урало-алтайской языковой общности совпадает с разделением гаплогруппы N1
на N1a, N1b и N1c. В среде гуннов также была значительная доля
сибирских субкладов гаплогруппы N и возможно легендарный вождь Аттила
был её представителем.
Первыми вождями принявшими участие в становлении венгерской империи
около 10 веков назад также были N1c1, угорский субклад L1034 что
подтвердило тестирование дДНК.

В Европе гаплогруппа N в
виде европейского субклада N1c1 чрезвычайно распространена вокруг всего
Балтийского моря, достигая максимальных частот в восточной Балтике, при
этом для южно-балтийского региона и южной Скандинавии характерен
субклад N1c1 L550.
Вероятно с участием гаплогруппы N1c1-L550 развилось мореходство на
Балтийском море, а путь из "Варяг в греки" с приходом Рюрика заложил
основу централизованного государства Русь.
По результатам тестирования документально-подтвержденных потомков установлено, что знаменитыми представителями рода являются:
основатель государственности на Руси -
Рюрик (Мономашичи:
Вадбольский, Лобанов-Ростовский, Хилков, Гагарин, Шаховский, Кропоткин,
Ржевский, Путянин, Татищев; Ольговичи: Масальский, Пузына), князь
литовский
Гедимин (Трубецкой, Голицин, Хованский), князь литовский
Гедрус (династия Довспрунгов)(и
его потомки Гедройцы), а также большое кол-во дворянских и купеческих
семей Речи Посполитой, Великого Княжества Литовского и Руси.
Гаплогруппа I1
Предполагается, что гаплогруппа I (общий предок для гаплогрупп
I1 и I2) образовалась в палеолите в Европе еще до последнего ледникового
максимума, и вероятно, родиной являлись Балканы или Пиренеи, где в
течении последнего ледникового периода находилась гаплогруппа I.
Затем, после отступления ледников, часть мезолитических
охотников-собирателей распространилась до севера Европы - гаплогруппа
I1.
Большинство современных носителей гаплогруппы I1 говорят
на германских языках, являющихся ветвью индоевропейского языкового
древа, однако до культурно-языкового влияния индо-европейцев данная
гаплогруппа предположительно говорила на другом, палеоевропейском
языке.
Возможно, что часть лексики появилась в германских языках от догерманского субстрата ассимилированных носителей гаплогруппы I1.
Все современные представители гаплогруппы I1 происходят от мужчины, жившего не более 5 тыс. лет назад.
Частота I1 в мире:
Шведы — 35,6%
Финны — 28,3%
Датчане — 32,8%
Норвежцы — 31,1%
Немцы — 15,2%
Эстонцы — 14,8%
Англичане — 14,4%
Французы — 9,8%
Русские — 7,6%
Украинцы — 4,9%
Ирландцы — 6,0%
Чехи — 8,5%
|
 Распространение гаплогруппы I1
Распространение гаплогруппы I1
|
Вопрос появления гаплогруппы в
России остается открытым. Возможны разные варианты: от потомков пленных
шведов и германских колонистов, до ассимилированных славянами остготов,
проживавших в Крыму с IV века н.э. Но большая часть I1 к русским
вероятно попала от финских народов- среди финнов это вторая по
распространенности гаплогруппа. Некоторые субклады гаплогруппы I1 могли
войти в состав праславянского, прагерманского или прафинского этносов
еще на самой заре их формирования.
| Популяция | Частота |
|---|
| Вологодская обл. | 18% |
| Архангельск | 14,2% |
| Рязанская обл. | 14% |
| Красноборск | 12,1% |
| Мокша | 12% |
| Вологда | 11,6% |
| Унжа (Костр.) | 11,5% |
| Кострома | 11,3% |
| Пензенская об. | 11% |
| Ивановская об. | 10% |
| Тамбовская об. | 10% |
| Карелы | 8,6% |
| Ливны (Орл.) | 8,2% |
| Тверь | 7,9% |
| Архангельск | 7,6% |
|
| Популяция | Частота |
|---|
| Чуваши | 7,5% |
| Архангельская об. | 7% |
| Брянская обл. | 7% |
| Остров (Пск.) | 6,7% |
| Костромская обл. | 6% |
| Псков | 5,4% |
| Русские Адыгеи | 5,1% |
| Тверская обл. | 5% |
| Курск | 5% |
| Казаки | 4,5% |
| Пристень (Курск.) | 4,4% |
| Кубанск. казаки ( | 4,4% |
| Казаки | 4,1% |
| Русские Башкирии | 4% |
|
| Популяция | Частота |
|---|
| Липецкая обл. | 4% |
| Коми | 3,6% |
| Белгород | 3,5% |
| Белгородская обл. | 3,5% |
| Воронеж | 3,1% |
| Новгородская обл. | 3% |
| Кашин (Тверская) | 2,7% |
| Вепсы | 2,6% |
| Смоленская обл. | 2% |
| Орловская обл. | 2% |
| Рославль (Смол.) | 1,9% |
| Смоленская обл. | 1,7% |
| Пинега (Арх.) | 0,9% |
| Татары | 0,8% |
|
Гаплогруппа I2
Гаплогруппа I2 зародилась в юго-восточной Европе более 15 тыс. лет
назад. Наивысшая её концентрация сегодня наблюдается в Балканских
странах: в Боснии-Герцеговине и южной Далмации (Хорватия), а также
составляет значительный % у всех южнославянских популяций.
Высокая плотность I2a в этих зонах даёт основания видеть в
Адриатическом регионе современной Хорватии и Динарских Альп природное
убежище для палеопопуляций-носителей I2a во время последнего ледникового
максимума.
Возможно представители гаплогруппы I2a стояли у истоков древнейших
культур Балканского полуострова, таких как: Чел Кладове, Лепенский Вир,
Гура Бачулуй, Неа Никомедия. На основе этого рода, возможно, сложились
иллирийские и фракийские племена. Ветвь I2a одной из первых заселила
Сардинию и на сегодняшний день данная гаплогруппа достигает там 40%. Тем
не менее все современные потомки в южной и восточной Европе относятся к
субкладу I2a2-Din, этому субкладу примерно 2500 лет, он возник на
границе Белоруссии и Украины, а на Балканы попал уже со славянскими
миграциями, либо незадолго до них.
Ветви гаплогруппы I2
I2a1
Гаплогруппа I2a1 (M26)
составляет около 40 % всех мужских линий восточной Сардинии. Кроме
того, она также обнаружена с низкой или умеренной частотой в Стране басков и на Иберийском полуострове, а также составила 1,6 % (1/64) в выборкеалбанцев, живущих в Македонии и 1,2 % (3/257) в выборке чехов. Возраст вариации YSTR для субклада M26 составляет 8,0±4.0 тыс. лет (Rootsi 2004).
Предполагается, что ветвь I2a выделилась из I2 около 11000 лет назад на Балканах, а ветвь I2a1 выделилась из I2a около 8000 лет назад на Сардинии или где-то на апенинском п-ве, в окрестностях Средиземного моря.
I2a2
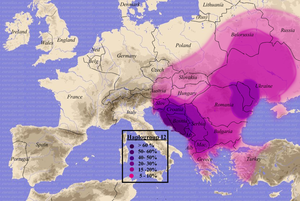
Распространение гаплогруппы I2a2
I2a2 (snp M423) типична для населения юго-восточной Европы. Наиболее высокая плотность — в Далмации (Хорватия) и Боснии-Герцеговине (>50 %). Предполагается, что в этой области на Динарском нагорье эта ветвь и произошла из I2a около 7500 лет назад. Чаще всего гаплогруппа I2a2 встречается средиславянских народов (особенно южнославянских), а также найдена среди румын,молдаван, венгров, южных литовцев, албанцев, греков, жителей северо-востокаИталии, в западной Анатолии и на Сардинии. Наиболее высокая частота гаплогруппы I2a2 среди
населения запада Балкан тем не менее не характеризует Балканы, как
прародину, потому что данный адриатический регион современной Хорватии
был перезаселен носителями данной гаплогруппы во время славянских
миграций. Ответвление snp P37.2 произошло около 10,7±4,8 тыс. лет назад
(Rootsi 2004). Возраст ветви snp P37.2 составляет 8,0±4,0 тысяч лет
(Rootsi 2004), а дляsnp M423 —- 8,8±3,6 тысяч лет (Underhill 2007).
Перичич считает, что распространение данной ветви произошло "не ранее
голоценового перехода и не позднее раннего неолита" (Pericic 2005).
Второй центр концентрации I2a2 расположен в нынешней Молдове и приблизительно совпадает с границами Трипольской культуры,
по-видимому, эта гаплогруппа являлась характерной для этой культуры, и с
приходом индоевропейцев влилась в их генофонд, не испытав такого
подавления, как другие ветви гаплогруппы I в центральной и западной Европе.
I2b1
Распространения гаплогруппы I2b1 совпадает с распространением гаплогруппы I1, за исключением Фенноскандии, что позволяет предположить её наличие как минимум в одном из палеолитических убежищ, где также находилась гаплогруппа I1. Отсутствие I2b1 в Фенноскандии может быть связано с тем, что на гаплогруппу I2b1 в наиболее ранних поселениях региона повлиял «эффект основателя» и генетический дрейф из-за её редкости, поскольку гаплогруппа I2b1 составляет менее 10 % от всего Y-хромосомного разнообразия населения на указанной территории за пределами Нижней Саксонии.
Распространение гаплогрупп I1 и I2b1 относительно хорошо коррелирует с историческими границами распространения носителей германских языков, однако первоначально данные линии говорили на каком-либо из палеоевропейских языков. Гаплогруппа I2b1 обнаружена
более чем у 4 % населения лишь в Германии, Нидерландах, Бельгии, Дании,
Англии (кроме Уэльса и Корнуолла), Шотландии, на южной оконечности
Швеции и Норвегии, а также в провинциях Нормандия, Мэн, Анжу и Перш на северо-западе Франции, в Провансе на
юго-востоке Франции, в исторических областях Италии — Тоскане, Умбрии и
Лациуме; а также в Молдавии, в Рязанской области и в Мордовии. В ходе
исследования дДНК региона севера Центральной Европы удалось установить,
что еще в 1 тыс. до нашей эры были этногруппы с 80% I2b1. Представляется
достаточно вероятным, что присутствие I1 и I2b1 в нынешних Франции,
Англии и Италии, а также в восточной Европе связано уже с кельтской и
германской экспансией, а в доиндоевропейское время эти гаплогруппы
концентрировались лишь в северной Европе. Одна из ветвей гаплогруппы I2b1, а именно I2b1a (snp
M284), обнаружена почти исключительно у населения Великобритании, что
может говорить о его длительной истории существования на Британских
островах. Интересно, что с небольшой частотой гаплогруппыI1 и I2b встречаются на территории исторических регионов Вифиния и Галатия в современной Турции, куда их могли принести кельты, мигрировавшие туда по приглашению Никомеда I Вифинского.
Гаплогруппа I2b1 также встречается примерно у 1 % населения Сардинии.
Предполагается, что I2b выделяется из I2 в центральной Европе, вблизи границы медленно отступающего ледника около 13 тыс. лет назад, I2b1 — из I2b ещё севернее, на территории нынешней Германии, около 9 тыс. лет назад. Специфическая британская ветвь I2b1a выделилась из I2b1 приблизительно 3 тысячелетия назад.
I2b2
Гаплогруппа I2b2 была обнаружена в скелетных останках, найденных в пещере Лихтенштейн — археологическом памятнике Бронзового века в центральной Германии, где также были найдены артефакты культуры Полей погребальных урн.
Из 19 мужских останков пещеры у 13 была обнаружена гаплогруппа I2b2, у
одного — R1b, и у двоих — R1a. Предположительно, пещера находилась в
эпицентре тогдашнего распространения гаплогруппы I2b.
Гаплогруппа E и E1b1b1
Гаплогруппа E1b1b1 (snp М35) объединяет около 5% всех мужчин на Земле и
насчитывает около 700 поколений до общего предка. Родоначальник
гаплогруппы E1b1b1 родился примерно 15 тысяч лет назад в Восточной
Африке (возможно, в Эфиопии).
В течение нескольких тысячелетий носители этой гаплогруппы жили на своей
исторической родине в Эфиопии и занимались охотой и собирательством. По
расовой принадлежности первоначально E и E1b1b1 были негроидными, но в
дальнейшем после миграций на север, хамиты относились к Кушитской
большой ветви Западного расового ствола и говорили на ностратическом или
афразийском праязыке. Согласно теории Дьяконова-Бендера, в Эфиопии
хамито-семитский праязык выделился из ностратического языка примерно 14
тыс. лет назад.
Примерно 13 тыс. лет назад климат на Земле стал меняться, и не в лучшую
сторону. Закончилась эпоха тепла и высокой влажности. Наступил
длительный период холодного и сухого климата. Вероятно, именно эти
изменения климата способствовали тому, что племена Восточной Африки,
принадлежащие преимущественно гаплогруппе E1b1b1, начали своё
движение из Эфиопии на север, в районы более благоприятные для жизни: в
Нубию, Египет и на Ближний Восток. В неолите род E1b1b1 распространился в
район Средиземноморья и в Южную Африку. Это расселение способствовало
обособлению отдельных групп E1b1b1. Появились отдельные народы, имеющие
свой язык и культуру: египтяне, берберы, ливийцы, кушиты, эфиопы,
химьяриты, ханаанеи и южно-африканские скотоводы. У мужчин этих новых
народов в Y-хромосоме появлялись новые SNP-мутации, которые они
передавали своим потомкам.
Таким образом, в роду E1b1b1-М35 появились субклады:
1. E1b1b1а (snp М78). Древние египтяне и их потомки, в том числе в
Европе: микенцы, македонцы, эпироты, отчасти ливийцы и нубийцы.
2. E1b1b1b (snp М81). Берберы. Потомки мавров в Европе.
3. E1b1b1с (snp М123). Потомки ханаанеев.
4. E1b1b1d (snp M281). Южные эфиопы (оромо).
5. E1b1b1e (snp V6). Северные эфиопы (амхара)
6. E1b1b1f (snp P72). Танзанийцы или эфиопы.
7. E1b1b1g (snp M293). Танзанийцы (датог, сандаве) и намибийцы (кхое).
Гаплогруппа E1b1b1а (snp М78) – основная гаплогруппа древних египтян.
Общий предок жил 11-12 тысяч лет назад. Род E1b1b1а (snp М78) стоял у истоков древнеегипетской цивилизации.
В бронзовом веке египтяне или их потомки переселились на Балканы. В
настоящее время гаплогруппа E1b1b1а наиболее распространена среди
албанцев и греков, и представлена балканскими субкладами:
E1b1b1a2 (snp V13) - потомки микенцев, македнов и эпирцев и
E1b1b1a5 (snp M521) возможно, потомки ионийцев.
Кроме вышеупомянутых двух субкладов, в гаплогруппе E1b1b1а выделяют ещё три:
E1b1b1a1 (snp V12) - потомки южных египтян
E1b1b1a3(snp V22) - потомки северных египтян и
E1b1b1a4 (snp V65) - ливийцы и марокканские берберы.
Потомки библейского Мицраима внесли огромный вклад в мировую историю,
искусство, науку и религию. Возможно именно представители гаплогруппы
E1b1b1а вывели первые агрокультуры, изобрели одну из самых ранних
письменностей, основали одно из величественных государств на Земле –
Древний Египет.
Первые E1b1b1 (V13) появились в Южной Европе уже 7 тыс. лет назад согласно дДНК.
Потомками древних египтян были братья Райт – создатели первого в мире
самолёта, способного к управляемому полёту, португальский навигатор и
исследователь Западной Африки Жоан Афонсу де Авейру, вице-президент США
Джон Колдвелл Кэлхоун и много других выдающихся людей.
Гаплогруппа E1b1a встречается почти исключительно среди жителей
западной, центральной и южной Африки. Это единственная Y-гаплогруппа,
являющаяся общей для всей субсахарской Африки, а также для потомков
африканских рабов в Америке и на Карибских островах. В других местах она
встречается с исчезающе малой частотой, и обычно её наличие объясняется
работорговлей, которую вели арабы в средние века.
Гаплогруппы E1 и E2 являются самыми распространенными в Африке
Гаплогруппа G
Гаплогруппа G возникла более 20 000 лет назад вероятно еще до
начала последнего ледникового периода в регионе современного Ирана.
Данная гаплогруппа, наряду с гаплогруппами J2a, J2b, J1 вероятно были
одними из первых людей, принявших участие в неолитической революции и
распространении земледелия и скотоводства сначала в регионе Ближнего
Востока в междуречье Тигра и Евфрата, а затем и в Южной Европе на
западе, в Египте на юге, и в Иране на востоке. Демографический взрыв гаплогруппы G был обеспечен огромными преимуществами неолитической революции.
Контроль над производством и распределением растительного продовольствия
привел к появлению цивилизации, централизованного управления. Вместо
маленьких кланов охотников-собирателей, перемещавшихся за стадами диких
животных, появились крупные оседлые сообщества земледельцев и скотоводов
с невиданной ранее сложной системой общественных отношений, социальной
лестницей, заложенной в основу иерархии. Переход человечества к
земледелию привел к появлению торговли, письменности, астрономических
календарей и возникновению крупных городов - конгломератов. Вместе с
распространением земледелия начали перемещаться c Ближнего востока и
предки гаплогруппы, так G попали в Турцию, на Балканы и на Кавказ, где в
настоящее время наблюдается самая высокая в мире плотность гаплогруппы.
Гаплогруппа G имеет множество субкладов, которые имеют свою древнюю и
занимательную историю.
Распространение
Кавказ
В настоящее время из довольно крупных народов, гаплогруппа G с наибольшей частотой встречается среди осетин (обнаружена у 68 % осетинских мужчин), в Дигорском и Алагирском районах Северной Осетии — до 76 %.
Из небольших популяций, очень высока частота встречаемости гаплогруппы G среди шапсугов (субклад G2a3b-P303) иказахского рода маджар — около 80 %.
Славяне
Как сказано выше, гаплогруппа G обнаружена у половины терских казаков.
Помимо них в Восточной Европе гаплогруппа G наиболее распространена на юге России и восточной Украине (не
более 4 %), что по некоторым исследованиям является результатом
включения традиционно обитавших там алан в состав пришедшего позднее
восточнославянского населения (аланы и восточные славяне).
В целом, среди русских и украинцев частота встречаемости гаплогруппы G — 1-2 %. Также с небольшой частотой (4-5 %) G встречается среди чехов и хорватов.
Европа
В остальной Европе гаплогруппа G
встречается редко (менее 4 % в целом по континенту) и представлена в
центральной её части отдельной подгруппой G2c. Проникновение этой
подгруппы относительно недавнее (менее 1000 лет назад) и связывается с
расселением евреев-ашкенази, у которых она встречается приблизительно в 8 % случаев.
Азия
Гаплогруппа J1
Частота гаплогруппы J1, определяемой маркером M267, резко падает на границах арабоговорящих стран и Дагестана с другими странами, такими как Иран (10.5%) и Турция (9%).
Аравийское плато
Гаплогруппа J1c3d, определяемая маркером P58, наиболее часта в Йемене (76%), Саудовской Аравии (64%) и Катаре (58%), также эта группа является обычной для Негевских бедуинов (62%). Среди других арабов она также очень распространена, особенно живущих в Леванте, т.е. палестинцев (38,4%), в Сирии (30%) и Ливане (25%). Среди еврейского населения J1c3dсоставляет 30% йеменских евреев, 20,0% ашкенази и 12% сефардов.
Предположительно, этому роду принадлежал легендарный Авраам.
Дагестан
Северная Африка
В Северной Африке (на основании характерности YCAIIa22-YCAIIb22 среди алжирцев 35,0% и тунисцев 31%), J1 впервые попала в Эфиопию с распространением семитоязычных племён (Эритрея 11%, Эфиопия 9%, Эфиопия-Амхара (33,3%). J1 также с высокой частотой регистрируется в северных частях Судана (J-12f2(xJ2-M172): суданские арабы 45%, нубийцы 41%, копты39%, беджа 36%), и с меньшей частотой присутствует в районе Darfur (J-12f2(xJ2-M172): Masalit 6%, Fur 6%). Гаплогруппа J1может быть найдена по крайней мере у 20% египетских мужчин.
Европа
В целом частота J1 очень низка. Однако высокая плотность была зафиксирована в центральных Адриатических районах ИталииГоргано (англ.) (17,2%), Пескара (15%), в средиземноморской Паоле (11,1%), южносицилийской Рагузе (10,7%), на Крите (8,3%),Мальте (7,8%), Кипре (6,2%), в Греции (5,3%).
Гаплогруппа J2
Гаплогруппа J2 зародилась примерно 18000 лет назад на Ближнем
Востоке или Анатолии и затем распространилась на Балканы и во всем
Средиземноморье. Представители гаплогруппы J2 изобрели земледелие и
первыми стали одомашнивать животных, тем самым подарив миру
скотоводство. В центральную Европу некоторые представители гаплогруппы
J2 могли прибыть с одной из нескольких миграционных волн из Анатолии,
частично с греками, финикийцами, или с римскими легионерами и
поселенцами в начале нашей эры.
В Европе гаплогруппа J2 наиболее
распространена в Греции и Италии, где на eё долю приходится до 30 %, а
также в Турции и на Кавказе. На северном Кавказе в высокой концентрации у
ингушей (до 87%) и чеченцев (до 67%). В Индии с достаточно высоким
процентом (в среднем до 10%) встречается подгруппа J2b (snp M241).
Другая подгруппа J2a ограничена, главным образом, северо-западом
субконтинента. Вероятно часть из них попали туда с неолитическими
миграциями, а часть при индо-европейском вторжении.
Большое разнообразие J2 в турецком и южных европейских поселениях
объясняется исторической связью с Эгейской культурой и с Ближним
Востоком. Многие члены гаплогруппы J2 были ограничены прибрежными
средиземноморскими областями. Именно морская торговля вокруг
Средиземноморья, возможно, наравне с неолитической сельскохозяйственной
миграцией, способствовала распространению гаплогруппы J2 всюду в
средиземноморском мире...
Гаплогруппа C3
Генетический маркер "SNP M217", определяющий эту гаплогруппу возник 20
000 лет назад у человека который жил где-то в древних Восточноазиатских
поселениях. Он дал начало большому роду, который со временем распался
на племена и начал расселяться с территории современной Монголии. Это
расселение способствовало обособлению отдельных ветвей рода, среди
которых можно выделить две:
1) C3c (M48) – монгольские народы, тунгусо-маньчжурские народы, коренные
народы Восточной Сибири, тюркские народы Сибири и Центральной Азии. К
этому же роду относятся два таинственных изолированных народа – юкагиры и
айны, происхождение языка которых до сих пор остается спорным среди
учёных.
2) C3d (M407) – часть якутов и небольшая часть китайцев.
Генеалогии полагают, что это распространение во многом обязано
завоевательным походам Чингисхана в XII-XIII веках. В данной гаплогруппе
выделяется очень многочисленная и молодая ветвь, чей предок жил в эпоху
Чингисхана. Приблизительно до 16 миллионов мужчин по некоторым
подсчетам, живущих в Центральной и Восточной Азии принадлежит этой
ветви. Профессор Оксфордской академии Брайан Сайкс объявил
этот кластер родом Чингисхана, однако по данным российских
исследователей, это не совсем так, скорее всего, родоначальником данного
субклада является предок Чингисхана, возможно его прапрадед.
Гаплогруппа О3
Эта гаплогруппа возникла около 15 000 лет назад от мужчины, который,
вероятно, родился на территории современного Китая. Более 80% китайцев
являются потомками этого человека по мужской линии.
Взрывной популяционный рост гаплогруппы О3 связан с неолитической
революцией в Восточной Азии - выращиванием риса. Имея секрет технологии
рисоводства позволявшего прокормить большую численность населения, гаплогруппа О3 сумела
подавить окружающие популяции и вырвалась в родовые лидеры восточной
Азии. Этот демографический рывок произошел примерно в то же самое время,
что и аграрная зерновая революция в районе Плодородного Полумесяца на
Ближнем Востоке. При археологических раскопках в северном Китае было
обнаружено просо, начало распространения которого связывают с
междуречьем Тигра и Евфрата.
4000 лет назад рисоводство вместе с представителями гаплогруппы О3
достигло островов Индонезии Борнео и Суматры, и сегодня носители O3
обнаруживаются вплоть до отдаленной Полинезии.
Деятели генеалогии принадлежащие гаплогруппе O3
Гаплогруппа Q
Гаплогруппа Q происходит от человека, родившегося в Сибири. Его
потомки Q1a-M3 были первыми исследователями Американского континента.
Несмотря на холодные температуры, часть гаплогруппы Q постепенно
продвигалась через свободную ото льда тундру с юга Сибири на
Северо-восток. Приблизительно 15 000 лет назад, они, дойдя до Чукотки,
сумели перебраться на соседний континент. Как им это удалось до сих пор
остается спорным вопросом. По одной из версий огромные массы воды в ту
пору были аккумулированы в леднике, и уровень мирового океана был на 100
метров ниже, чем сейчас. Следовательно, Сибирь и Аляска были соединены
перешейком, через который гаплогруппа Q благополучно перебралась в
Америку и продолжила свой путь на юг, создавая по пути великие
американские цивилизации Инков и Ацтеков.

Род Ашина, возможно происходил из ветви оставшейся в Сибири части
гаплогруппы Q, и сумел объединить под своим началом разнородные племена
(R1a-Z93, N1), положил начало созданию Тюркского каганата. По некоторым
данным, тюркский язык мог и быть языком рода Ашина, который восприняли
все объединенные в Каганате рода. После распада Тюркского каганата
правящая верхушка перебралась в отдаленную провинцию - в Хазарский
каганат, где и правила до 10 века. ДНК-генеалоги получили гаплотип
предполагаемых потомков рода Ашина - оказалось Q1b, и сейчас потомки
рода имеют свой ДНК - проект. Среди участников данного ДНК-генеалогического проекта имеются некоторые известные москвичи.
Гаплогруппы: описание древних родов и связь с археологическими культурами












































































