************************************************************
воскресенье, 3 мая 2015 г.
Гаплогруппы: описание древних родов и известные представители гаплогрупп
 Гаплогруппа
Гаплогруппа

(в
популяционной генетике человека, науке, изучающей генетическую историю
человечества) — большая группа схожих гаплотипов, которые являются рядом
аллелей на нерекомбинируемых участках Y-хромосомы. Галпогруппы делятся
на Y-хромосомные (Y-ДНК) и митохондриальные (мт-ДНК). Y-ДНК является
прямой отцовской линией, т.е сын, отец, дед и т.д., а мт-ДНК прямой
материнской линией, то есть дочь, мать, бабушка, прабабушка и так далее.
Термин «гаплогруппа» широко применяется в генетической ДНК генеалогии.
Гаплогруппа R1a










Гаплогруппа R1a1 насчитывает около 300 миллионов мужчин. Первый общий предок современных носителей R1a1 жил около 300 поколений назад.
Распространение гаплогруппы R1a :
В процентах указана доля R1a от общей численности этноса
- Русские 48%
- Поляки 56%
- Украинцы 54%
- Белорусы 51%
- Чехи 34%
- Киргизы 63%
- Шорцы 56%
- Алтайцы 54%
- Чуваши 31,5%
- Таджики 53%
- Панджабцы 54% (Пакистан-Индия)
- Индия в целом 30%, верхние касты 43%
Экскурс в Древнюю историю гаплогруппы R1a
Гаплогруппа R1a1 возникла
около 15 000 лет назад в Азии и в дальнейшем распалась на несколько
субкладов, или как их еще называют, дочерних гаплогрупп. Мы рассмотрим
основные из них - это Z283 и Z93. R1a1-Z93 - это азиатский маркер,
характерный для тюрков, евреев, индийцев. С участием гаплогруппы
R1a1-Z93 изобрели колесо в степи, сконструировали первые повозки и
приручили лошадь. Это были культуры Андроновского круга. Гаплогруппа
быстро освоила всю полосу евразийских степей от Каспия до Забайкалья,
распавшись на множество различных племен с разными этнокультурными
особенностями.
R1a1-Z283 - это европейский маркер и характерен по большей части для
славян, но не только, свои отдельные субклады имеются и у скандинавов,
англичан. В целом на сегодняшний день древняя гаплогруппа R1a1 наиболее характерна для славянских, тюркских и индийских этносов.

Раскопки "Страны городов" на Южном Урале подтвердили, что уже примерно
4000 лет назад в укрепленном поселении Аркаим имелись помещения личного и
общественного пользования, жилые и мастерские. В некоторых помещениях
обнаружены не только гончарные мастерские, но и металлургическое
производство.
В ходе раскопок было вскрыто около 8000 кв. м площади поселения
(примерно половина), вторая часть была исследована с помощью
археомагнитных методов. Тем самым была полностью установлена планировка
памятника. Здесь же был впервые в Зауралье применён метод реконструкции,
и Л.Л. Гуревичем сделаны рисунки возможного вида поселения. R1a1-Z93
вероятно была одной из основных гаплогрупп в Аркаиме.
Многие племена, вступая в контакт с этими общинами постепенно переходили
на их язык. В настоящее время почти вся Европа говорит на
индоевропейских языках, в то время как гаплогруппа R1b более специфична западной Европе, а R1a - восточной Европе. В странах находящихся ближе к центральной Европе имеются обе эти гаплогруппы. Так гаплогруппа R1a занимает
около 30% в населении Норвегии, и небольшой процент (не более 15%) в
Восточной Германии - остатки прямых Y-линий некогда ассимилированных
немцами полабских славян.

Во втором тысячелетии до нашей эры предположительно из-за климатических
изменений часть R1a1 (субклад Z93 и др. гаплогруппы Средней Азии) начали
мигрировать на юг и восток за пределы степи, часть (субклад L657) пошла
в сторону Индии и, влившись в местные племена, приняла участие в
создании кастового общества. Те далекие события описаны в древнейшем
литературном источнике человечества –"Ригведе".
Другая часть стала продвигаться в ближневосточном направлении. На территории современной Турции предположительно они основали Хеттское государство,
успешно конкурировавшее и с древним Египтом. Хетты строили города, но
не смогли прославиться сооружением огромных пирамид, так как в отличие
от Египта Хеттское общество было обществом свободных людей, и им
были чужды идеи использования подневольного труда. Хеттское государство
исчезло внезапно, его смела мощная волна варварских племен известных
как "народы моря". В середине прошлого века археологи нашли богатейшую
библиотеку глиняных табличек с хеттскими текстами, язык оказался
относящимся к индо-европейской группе языков. Так мы обрели детальные
знания о первом государстве, часть мужских линий которого
предположительно состояла из гаплогруппы R1a1-Z93.
Славянские субклады гаплогруппы R1a1-Z283 образуют свой кластер гаплотипов, который совершенно не родственен ни западно-европейским субкладам гаплогруппы R1a, ни индо-иранским и разделение европейских носителей R1a1-Z283 с азиатскими R1a1-Z93 произошло примерно 6 500 лет назад.

В октябре 539 года (до нашей эры) иранское племя Персов захватило Вавилон, вождь персов Кир решил
не уходить, а серьезно обосноваться в захваченном городе. Впоследствии
Киру удалось значительно расширить свои владения, так возникла великая
Персидская Империя, просуществовавшая дольше всех империй в мире - 1190
лет! В 651 году нашей эры ослабленная междоусобицей Персия пала под
натиском арабов, и это привело к изменению гаплогруппного состава
населения. Сейчас на обширных территориях, занимаемых некогда иранским
государством, практически не осталосьгаплогруппы R1a, по иронии судьбы Иран, остался без гаплогруппы, внесшей большой вклад на этапе становления этого государства.
Три мировые религии связаны с индо-ариями - Индуизм, Буддизм и Зороастризм.
Большинство народов состоит из множества гаплогрупп, и не имеется рода,
который бы доминировал над остальными. Связь между гаплогруппой и
внешностью человека также отсутствует и как видно многие представители гаплогруппы R1a1 относятся даже к разным расам. Многим R1a1-Z93
характерны монголоидные черты (киргизы, алтайцы, хотоны и т.д.), в то
время как носители R1a1-Z283 имеют в большинстве европейский вид
(поляки, русские, белорусы и т.д.). Большое количество финских племен
имеют высокие проценты гаплогруппы R1a1, часть из которых были ассимилированы с приходом славянских колонистов 9 века.
Достижения, к которым возможно имеют отношение R1a1:
Колесо, повозки, приручение лошади, металлургия, брюки, сапоги, платья,
первый в мире мощеный "автобан" протяженностью более 1000 км со
станциями "заправки" - замены лошадей, и много чего другого.
В небольшой статье трудно рассказать всю историю первых индо-европейцев,
можно лишь некоторыми историческими фрагментами пробудить интерес к
истории древних предков славян. Наберите в поисковике слова индо-арии, тюрки, славяне,скифы, сарматы, Персия, и Вы окунетесь в увлекательное путешествие по славной истории индо-европейских и славянских народов.
Древо гаплогруппы.
До 2007 года никто не осуществляли детальных реконструкций родов, никому
в голову не пришла эта идея, да и решить такую грандиозную задачу не
было возможности. Многие популяционные генетики работали с небольшими
выборками коротких 6-маркерных гаплотипов, позволяющих получать общие
генографические представления о распределении гаплогрупп.
 В
2009 году профессиональный популяционный генетик задался целью
построить детальное генеалогическое древо этой гаплогруппы. Столкнувшись
с рядом проблем, например, расчет обычными методами крупных выборок
сверх длинных гаплотипов был невозможен из-за астрономического
количества операций, ни один компьютер был не в состоянии перебрать
нужное количество комбинаций, но благодаря находчивости и желанию
построить древо своей гаплогруппы проблемы эти удалось преодолеть.
В
2009 году профессиональный популяционный генетик задался целью
построить детальное генеалогическое древо этой гаплогруппы. Столкнувшись
с рядом проблем, например, расчет обычными методами крупных выборок
сверх длинных гаплотипов был невозможен из-за астрономического
количества операций, ни один компьютер был не в состоянии перебрать
нужное количество комбинаций, но благодаря находчивости и желанию
построить древо своей гаплогруппы проблемы эти удалось преодолеть.
После R1a1 многие гаплогруппы приступили к созданию своих древ.
Сами гаплогруппы не несут генетической информации, т.к. генетическая
информация находиться в аутосомах- первых 22 пар хромосом. Посмотреть
распространение генетических компонент в Европе можно здесь. Гаплогруппы являются всего лишь метками давно минувших дней, на заре формирования современных народов.
Гаплогруппа R1b









Гаплогруппа
R1b – параллельный субклад для гаплогруппы R1a. Родоначальник
гаплогруппы R1b родился около 16 000 лет назад в центральной Азии от
родительского рода R1. Примерно 10 000 лет назад гаплогруппа R1b
распалась на несколько субкладов, которые начали расходиться в разных
направлениях. Восточную ветвь - субклад R1b-M73 некоторые ученые
связывают с древними тохарами, которые приняли участие в этногенезе
такого народа, как современные уйгуры.

Продвижение гаплогруппы R1b на
запад в Европу вероятно происходило в несколько этапов. Часть может
быть связана с неолитическими миграциями из Малой Азии и Закавказья, а
часть с миграциями постнеолитическими и распространением археологической
культуры Колоколовидных Кубков. Существует версия и о миграции по
Североафриканскому побережью к Гибралтарскому проливу, с дальнейшей
переправкой на Пиренеи в виде археологической культуры Колоколовидных
кубков - но в данной гипотезе слишком много натяжек. В любом случае
большинство европейских представителей гаплогруппы R1b имеют снип P312,
который определенно произошел уже в Европе.
После того, как египетские ученые провели анализ мумии Тутанхамона, было обнаружено, что фараон оказался представителем гаплогруппы R1b.
Сейчас основная масса представителейгаплогруппы R1b1a2 проживает в Западной Европе, где гаплогруппа R1b1a2 является
основной гаплогруппой. В России только башкирский народ имеет большой
процент этой гаплогруппы. В русском народе гаплогруппа R1b составляет не
более 5%. В Петровскую и Екатерининскую эпоху проводилась
государственная политика массового привлечения иностранных специалистов
из Германии и остальной Европы, многие русские R1b являются их
потомками. Также какая-то часть могла войти в русский этнос с Востока -
это прежде всего субклад R1b-M73. Некоторые R1b-L23 могут быть
мигрантами с Кавказа, куда они попали из Закавказья и Передней Азии.
Европа
Современная концентрация гаплогруппы R1b максимальна
на территориях миграционных путей кельтов и германцев: в южной Англии
около 70%, в северной и западной Англии, Испании, Франции, Уэльсе,
Шотландии, Ирландии — до 90 % и более. А так же, например, у басков — 88,1 %, испанцев — 70 %, итальянцев — 40 %, бельгийцев — 63 %, немцев — 39 %, норвежцев — 25,9 % и других.
В Восточной Европе гаплогруппа R1b встречается намного реже. У чехов и словаков — 35,6 %, латышей — 10 %, венгров — 12,1 %, эстонцев — 6 %, поляков — 10,2 %-16,4 %, литовцев — 5 %, белорусов — 4,2 %, русских — от 1,3 % до 14,1 %,украинцев — от 2 % до 11,1 %.
На Балканах — у греков — от 13,5 % до 22,8 %, словенцев — 21 %, албанцев — 17,6 %, болгар — 17 %, хорватов — 15,7 %,румын — 13 %, сербов — 10,6 %, герцеговинцев — 3,6 %, боснийцев — 1,4 %.
Азия
На Южном Урале значительно распространена у башкир — около 43 %.
В Центральной Азии обнаружена, в частности, у туркменов — 36,7 %, узбеков — 9,8 %, татар — 8,7 %, казахов — 5,6 %,уйгуров — от 8,2 % до 19,4 %
Африка
У алжирских арабов из Орана — 10,8 %, тунисских арабов — 7 %, алжирских берберов — 5,8 %, в Марокко — около 2,5 %, в субсахарской Африке распространена в Камеруне — около 95 % (субклад R1b-V88).
Гаплогруппа N












Гаплогруппа N возникла около 15 000 лет назад на юго-западе современного
Китая (провинция Юннань), и около 10 000 лет назад начала свой путь
движения в сторону Байкала, Саян и Алтая. На Байкале часть
представителей гаплогруппы N осталась, часть направилась на северо-запад
в сторону Балтики, а другая часть в сторону Дальнего Востока. Пройдя
через всю Сибирь, Урал и Русскую равнину, племена достигли побережья
Балтийского моря около 4 000 лет назад предположительно в рамках
археологической культуры Ямочно-Гребенчатой Керамики.

На всем пути продвижения на базе рода формировались разные этносы, по
большей части финно-угорские, тюркские, славянские и балтские (снип
L1025). На сегодняшний день максимальные частоты N1c1 обнаружены у
финнов и якутов, у которых гаплогруппа N1с1 достигает 80%, а
также балты (латыши и литовцы) - у них N1c1 достигает 47%. Современные
народы у которых преобладает общая гаплогруппа зачастую совершенно
различны антропологически, т.к. имеют разные субклады, разделенные около
5-7 тыс. лет назад, это объясняется просто: осваивать новые земли
отправлялись здоровые, молодые мужчины, а старики, женщины и дети
оставались на месте. Молодые войны берут жен из местных племен и таким
образом, каждое новое поколение, произошедшее от европейских женщин,
становилось все более и более европейским по внешнему виду.
В русском этносе гаплогруппа N занимает почетное второе место по распространенности. Наличие гаплогруппы N1с1 в
генофонде русских объясняется с одной стороны наличием субклада N1c1
L550 у славян (включая западных) а также ассимиляцией некоторых
финно-угорских народов славянскими колонистами 9 века; наличие
гаплогруппы N1c1 у поволжских народов тоже достигает высоких процентов.
Такие народы как мурома, полностью растворились в русских, но их мужские
линии продолжились в русском и других этносах до настоящего времени.

Гаплогруппа N1 прослеживается также в южном Китае, Бирме, Тайланде, Камбодже, Японии, Тайване и Корее.
Результаты дДНК подтвердили наличие N1 в останках культуры Яншао.
Возникновение тюркских языков многие исследователи также связывают с
гаплогруппой N1, возможно N1b. Реконструкция времени разделения
урало-алтайской языковой общности совпадает с разделением гаплогруппы N1
на N1a, N1b и N1c. В среде гуннов также была значительная доля
сибирских субкладов гаплогруппы N и возможно легендарный вождь Аттила
был её представителем.
Первыми вождями принявшими участие в становлении венгерской империи
около 10 веков назад также были N1c1, угорский субклад L1034 что
подтвердило тестирование дДНК.

В Европе гаплогруппа N в
виде европейского субклада N1c1 чрезвычайно распространена вокруг всего
Балтийского моря, достигая максимальных частот в восточной Балтике, при
этом для южно-балтийского региона и южной Скандинавии характерен
субклад N1c1 L550.
Вероятно с участием гаплогруппы N1c1-L550 развилось мореходство на
Балтийском море, а путь из "Варяг в греки" с приходом Рюрика заложил
основу централизованного государства Русь.
По результатам тестирования документально-подтвержденных потомков установлено, что знаменитыми представителями рода являются: основатель государственности на Руси - Рюрик (Мономашичи:
Вадбольский, Лобанов-Ростовский, Хилков, Гагарин, Шаховский, Кропоткин,
Ржевский, Путянин, Татищев; Ольговичи: Масальский, Пузына), князь
литовский Гедимин (Трубецкой, Голицин, Хованский), князь литовский Гедрус (династия Довспрунгов)(и
его потомки Гедройцы), а также большое кол-во дворянских и купеческих
семей Речи Посполитой, Великого Княжества Литовского и Руси.
Гаплогруппа I1






Предполагается, что гаплогруппа I (общий предок для гаплогрупп
I1 и I2) образовалась в палеолите в Европе еще до последнего ледникового
максимума, и вероятно, родиной являлись Балканы или Пиренеи, где в
течении последнего ледникового периода находилась гаплогруппа I.
Затем, после отступления ледников, часть мезолитических
охотников-собирателей распространилась до севера Европы - гаплогруппа
I1.
Большинство современных носителей гаплогруппы I1 говорят
на германских языках, являющихся ветвью индоевропейского языкового
древа, однако до культурно-языкового влияния индо-европейцев данная
гаплогруппа предположительно говорила на другом, палеоевропейском
языке.
Возможно, что часть лексики появилась в германских языках от догерманского субстрата ассимилированных носителей гаплогруппы I1.
Все современные представители гаплогруппы I1 происходят от мужчины, жившего не более 5 тыс. лет назад.
Частота I1 в мире: Шведы — 35,6% Финны — 28,3% Датчане — 32,8% Норвежцы — 31,1% Немцы — 15,2% Эстонцы — 14,8% Англичане — 14,4% Французы — 9,8% Русские — 7,6% Украинцы — 4,9% Ирландцы — 6,0% Чехи — 8,5% |  Распространение гаплогруппы I1 |
Вопрос появления гаплогруппы в
России остается открытым. Возможны разные варианты: от потомков пленных
шведов и германских колонистов, до ассимилированных славянами остготов,
проживавших в Крыму с IV века н.э. Но большая часть I1 к русским
вероятно попала от финских народов- среди финнов это вторая по
распространенности гаплогруппа. Некоторые субклады гаплогруппы I1 могли
войти в состав праславянского, прагерманского или прафинского этносов
еще на самой заре их формирования.
|
|
|
Гаплогруппа I2



Гаплогруппа I2 зародилась в юго-восточной Европе более 15 тыс. лет
назад. Наивысшая её концентрация сегодня наблюдается в Балканских
странах: в Боснии-Герцеговине и южной Далмации (Хорватия), а также
составляет значительный % у всех южнославянских популяций.

Высокая плотность I2a в этих зонах даёт основания видеть в
Адриатическом регионе современной Хорватии и Динарских Альп природное
убежище для палеопопуляций-носителей I2a во время последнего ледникового
максимума.
Возможно представители гаплогруппы I2a стояли у истоков древнейших
культур Балканского полуострова, таких как: Чел Кладове, Лепенский Вир,
Гура Бачулуй, Неа Никомедия. На основе этого рода, возможно, сложились
иллирийские и фракийские племена. Ветвь I2a одной из первых заселила
Сардинию и на сегодняшний день данная гаплогруппа достигает там 40%. Тем
не менее все современные потомки в южной и восточной Европе относятся к
субкладу I2a2-Din, этому субкладу примерно 2500 лет, он возник на
границе Белоруссии и Украины, а на Балканы попал уже со славянскими
миграциями, либо незадолго до них.
Ветви гаплогруппы I2
I2a1
Гаплогруппа I2a1 (M26)
составляет около 40 % всех мужских линий восточной Сардинии. Кроме
того, она также обнаружена с низкой или умеренной частотой в Стране басков и на Иберийском полуострове, а также составила 1,6 % (1/64) в выборкеалбанцев, живущих в Македонии и 1,2 % (3/257) в выборке чехов. Возраст вариации YSTR для субклада M26 составляет 8,0±4.0 тыс. лет (Rootsi 2004).
Предполагается, что ветвь I2a выделилась из I2 около 11000 лет назад на Балканах, а ветвь I2a1 выделилась из I2a около 8000 лет назад на Сардинии или где-то на апенинском п-ве, в окрестностях Средиземного моря.
I2a2
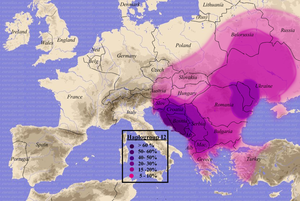
Распространение гаплогруппы I2a2
I2a2 (snp M423) типична для населения юго-восточной Европы. Наиболее высокая плотность — в Далмации (Хорватия) и Боснии-Герцеговине (>50 %). Предполагается, что в этой области на Динарском нагорье эта ветвь и произошла из I2a около 7500 лет назад. Чаще всего гаплогруппа I2a2 встречается средиславянских народов (особенно южнославянских), а также найдена среди румын,молдаван, венгров, южных литовцев, албанцев, греков, жителей северо-востокаИталии, в западной Анатолии и на Сардинии. Наиболее высокая частота гаплогруппы I2a2 среди
населения запада Балкан тем не менее не характеризует Балканы, как
прародину, потому что данный адриатический регион современной Хорватии
был перезаселен носителями данной гаплогруппы во время славянских
миграций. Ответвление snp P37.2 произошло около 10,7±4,8 тыс. лет назад
(Rootsi 2004). Возраст ветви snp P37.2 составляет 8,0±4,0 тысяч лет
(Rootsi 2004), а дляsnp M423 —- 8,8±3,6 тысяч лет (Underhill 2007).
Перичич считает, что распространение данной ветви произошло "не ранее
голоценового перехода и не позднее раннего неолита" (Pericic 2005).
Второй центр концентрации I2a2 расположен в нынешней Молдове и приблизительно совпадает с границами Трипольской культуры,
по-видимому, эта гаплогруппа являлась характерной для этой культуры, и с
приходом индоевропейцев влилась в их генофонд, не испытав такого
подавления, как другие ветви гаплогруппы I в центральной и западной Европе.
I2b1
Распространения гаплогруппы I2b1 совпадает с распространением гаплогруппы I1, за исключением Фенноскандии, что позволяет предположить её наличие как минимум в одном из палеолитических убежищ, где также находилась гаплогруппа I1. Отсутствие I2b1 в Фенноскандии может быть связано с тем, что на гаплогруппу I2b1 в наиболее ранних поселениях региона повлиял «эффект основателя» и генетический дрейф из-за её редкости, поскольку гаплогруппа I2b1 составляет менее 10 % от всего Y-хромосомного разнообразия населения на указанной территории за пределами Нижней Саксонии.
Распространение гаплогрупп I1 и I2b1 относительно хорошо коррелирует с историческими границами распространения носителей германских языков, однако первоначально данные линии говорили на каком-либо из палеоевропейских языков. Гаплогруппа I2b1 обнаружена
более чем у 4 % населения лишь в Германии, Нидерландах, Бельгии, Дании,
Англии (кроме Уэльса и Корнуолла), Шотландии, на южной оконечности
Швеции и Норвегии, а также в провинциях Нормандия, Мэн, Анжу и Перш на северо-западе Франции, в Провансе на
юго-востоке Франции, в исторических областях Италии — Тоскане, Умбрии и
Лациуме; а также в Молдавии, в Рязанской области и в Мордовии. В ходе
исследования дДНК региона севера Центральной Европы удалось установить,
что еще в 1 тыс. до нашей эры были этногруппы с 80% I2b1. Представляется
достаточно вероятным, что присутствие I1 и I2b1 в нынешних Франции,
Англии и Италии, а также в восточной Европе связано уже с кельтской и
германской экспансией, а в доиндоевропейское время эти гаплогруппы
концентрировались лишь в северной Европе. Одна из ветвей гаплогруппы I2b1, а именно I2b1a (snp
M284), обнаружена почти исключительно у населения Великобритании, что
может говорить о его длительной истории существования на Британских
островах. Интересно, что с небольшой частотой гаплогруппыI1 и I2b встречаются на территории исторических регионов Вифиния и Галатия в современной Турции, куда их могли принести кельты, мигрировавшие туда по приглашению Никомеда I Вифинского.
Гаплогруппа I2b1 также встречается примерно у 1 % населения Сардинии.
Предполагается, что I2b выделяется из I2 в центральной Европе, вблизи границы медленно отступающего ледника около 13 тыс. лет назад, I2b1 — из I2b ещё севернее, на территории нынешней Германии, около 9 тыс. лет назад. Специфическая британская ветвь I2b1a выделилась из I2b1 приблизительно 3 тысячелетия назад.
I2b2
Гаплогруппа I2b2 была обнаружена в скелетных останках, найденных в пещере Лихтенштейн — археологическом памятнике Бронзового века в центральной Германии, где также были найдены артефакты культуры Полей погребальных урн.
Из 19 мужских останков пещеры у 13 была обнаружена гаплогруппа I2b2, у
одного — R1b, и у двоих — R1a. Предположительно, пещера находилась в
эпицентре тогдашнего распространения гаплогруппы I2b.
Гаплогруппа E и E1b1b1











Гаплогруппа E1b1b1 (snp М35) объединяет около 5% всех мужчин на Земле и
насчитывает около 700 поколений до общего предка. Родоначальник
гаплогруппы E1b1b1 родился примерно 15 тысяч лет назад в Восточной
Африке (возможно, в Эфиопии).
В течение нескольких тысячелетий носители этой гаплогруппы жили на своей
исторической родине в Эфиопии и занимались охотой и собирательством. По
расовой принадлежности первоначально E и E1b1b1 были негроидными, но в
дальнейшем после миграций на север, хамиты относились к Кушитской
большой ветви Западного расового ствола и говорили на ностратическом или
афразийском праязыке. Согласно теории Дьяконова-Бендера, в Эфиопии
хамито-семитский праязык выделился из ностратического языка примерно 14
тыс. лет назад.
Примерно 13 тыс. лет назад климат на Земле стал меняться, и не в лучшую
сторону. Закончилась эпоха тепла и высокой влажности. Наступил
длительный период холодного и сухого климата. Вероятно, именно эти
изменения климата способствовали тому, что племена Восточной Африки,
принадлежащие преимущественно гаплогруппе E1b1b1, начали своё
движение из Эфиопии на север, в районы более благоприятные для жизни: в
Нубию, Египет и на Ближний Восток. В неолите род E1b1b1 распространился в
район Средиземноморья и в Южную Африку. Это расселение способствовало
обособлению отдельных групп E1b1b1. Появились отдельные народы, имеющие
свой язык и культуру: египтяне, берберы, ливийцы, кушиты, эфиопы,
химьяриты, ханаанеи и южно-африканские скотоводы. У мужчин этих новых
народов в Y-хромосоме появлялись новые SNP-мутации, которые они
передавали своим потомкам.
Таким образом, в роду E1b1b1-М35 появились субклады:
1. E1b1b1а (snp М78). Древние египтяне и их потомки, в том числе в
Европе: микенцы, македонцы, эпироты, отчасти ливийцы и нубийцы.
2. E1b1b1b (snp М81). Берберы. Потомки мавров в Европе.
3. E1b1b1с (snp М123). Потомки ханаанеев.
4. E1b1b1d (snp M281). Южные эфиопы (оромо).
5. E1b1b1e (snp V6). Северные эфиопы (амхара)
6. E1b1b1f (snp P72). Танзанийцы или эфиопы.
7. E1b1b1g (snp M293). Танзанийцы (датог, сандаве) и намибийцы (кхое).


Гаплогруппа E1b1b1а (snp М78) – основная гаплогруппа древних египтян.
Общий предок жил 11-12 тысяч лет назад. Род E1b1b1а (snp М78) стоял у истоков древнеегипетской цивилизации.
В бронзовом веке египтяне или их потомки переселились на Балканы. В
настоящее время гаплогруппа E1b1b1а наиболее распространена среди
албанцев и греков, и представлена балканскими субкладами:
E1b1b1a2 (snp V13) - потомки микенцев, македнов и эпирцев и
E1b1b1a5 (snp M521) возможно, потомки ионийцев.
Кроме вышеупомянутых двух субкладов, в гаплогруппе E1b1b1а выделяют ещё три:
E1b1b1a1 (snp V12) - потомки южных египтян
E1b1b1a3(snp V22) - потомки северных египтян и
E1b1b1a4 (snp V65) - ливийцы и марокканские берберы.
Потомки библейского Мицраима внесли огромный вклад в мировую историю,
искусство, науку и религию. Возможно именно представители гаплогруппы
E1b1b1а вывели первые агрокультуры, изобрели одну из самых ранних
письменностей, основали одно из величественных государств на Земле –
Древний Египет.
Первые E1b1b1 (V13) появились в Южной Европе уже 7 тыс. лет назад согласно дДНК.
Потомками древних египтян были братья Райт – создатели первого в мире
самолёта, способного к управляемому полёту, португальский навигатор и
исследователь Западной Африки Жоан Афонсу де Авейру, вице-президент США
Джон Колдвелл Кэлхоун и много других выдающихся людей.
Гаплогруппа E1b1a встречается почти исключительно среди жителей
западной, центральной и южной Африки. Это единственная Y-гаплогруппа,
являющаяся общей для всей субсахарской Африки, а также для потомков
африканских рабов в Америке и на Карибских островах. В других местах она
встречается с исчезающе малой частотой, и обычно её наличие объясняется
работорговлей, которую вели арабы в средние века.
Гаплогруппы E1 и E2 являются самыми распространенными в Африке
Гаплогруппа G




Гаплогруппа G возникла более 20 000 лет назад вероятно еще до
начала последнего ледникового периода в регионе современного Ирана.
Данная гаплогруппа, наряду с гаплогруппами J2a, J2b, J1 вероятно были
одними из первых людей, принявших участие в неолитической революции и
распространении земледелия и скотоводства сначала в регионе Ближнего
Востока в междуречье Тигра и Евфрата, а затем и в Южной Европе на
западе, в Египте на юге, и в Иране на востоке. Демографический взрыв гаплогруппы G был обеспечен огромными преимуществами неолитической революции.
Контроль над производством и распределением растительного продовольствия
привел к появлению цивилизации, централизованного управления. Вместо
маленьких кланов охотников-собирателей, перемещавшихся за стадами диких
животных, появились крупные оседлые сообщества земледельцев и скотоводов
с невиданной ранее сложной системой общественных отношений, социальной
лестницей, заложенной в основу иерархии. Переход человечества к
земледелию привел к появлению торговли, письменности, астрономических
календарей и возникновению крупных городов - конгломератов. Вместе с
распространением земледелия начали перемещаться c Ближнего востока и
предки гаплогруппы, так G попали в Турцию, на Балканы и на Кавказ, где в
настоящее время наблюдается самая высокая в мире плотность гаплогруппы.
Гаплогруппа G имеет множество субкладов, которые имеют свою древнюю и
занимательную историю.
Распространение
Кавказ
В настоящее время из довольно крупных народов, гаплогруппа G с наибольшей частотой встречается среди осетин (обнаружена у 68 % осетинских мужчин), в Дигорском и Алагирском районах Северной Осетии — до 76 %.
Из небольших популяций, очень высока частота встречаемости гаплогруппы G среди шапсугов (субклад G2a3b-P303) иказахского рода маджар — около 80 %.
Затем, после маджар, шапсугов и осетин по частоте наибольшей распространенности данной гаплогруппы идут абхазы итерские казаки — 45 %. В Карачаево-Черкесии, Кабардино-Балкарии, Сванетии и Шида Картли(Грузия) — около 30 %. НаКавказе к востоку от Осетии — среди чеченцев, ингушей и народов Дагестана гаплогруппа G встречается значительно реже — с частотой до 5 %. Исключение составляют лезгины и аварцы с частотой около 13 % и 9 % соответственно.
Славяне
Как сказано выше, гаплогруппа G обнаружена у половины терских казаков.
Помимо них в Восточной Европе гаплогруппа G наиболее распространена на юге России и восточной Украине (не
более 4 %), что по некоторым исследованиям является результатом
включения традиционно обитавших там алан в состав пришедшего позднее
восточнославянского населения (аланы и восточные славяне).
В целом, среди русских и украинцев частота встречаемости гаплогруппы G — 1-2 %. Также с небольшой частотой (4-5 %) G встречается среди чехов и хорватов.
Европа
В других местах Европы гаплогруппа G довольно часто встречается в материковой Греции, северной Испании и Италии, наКрите, Сардинии и в Тироле (до 15 %). На юге Германии и в Венгрии встречается с частотой до 6 %.
В остальной Европе гаплогруппа G
встречается редко (менее 4 % в целом по континенту) и представлена в
центральной её части отдельной подгруппой G2c. Проникновение этой
подгруппы относительно недавнее (менее 1000 лет назад) и связывается с
расселением евреев-ашкенази, у которых она встречается приблизительно в 8 % случаев.
Азия
В странах Азии гаплогруппа G с частотой 10-20 % встречается у иранских персов, пуштунов (афганцев) и калашей. Также встречается у турок и армян с частотой 11 %, у иракских арабов с частотой около 10 % и у курдов. Менее распространена вПакистане, Индии (брахманы). Обнаружена у курейшитов. Самое большое скопление разновидностей гаплогруппы находится вЛиване (Финикия)
Гаплогруппа J1




Частота гаплогруппы J1, определяемой маркером M267, резко падает на границах арабоговорящих стран и Дагестана с другими странами, такими как Иран (10.5%) и Турция (9%).
Аравийское плато
Гаплогруппа J1c3d, определяемая маркером P58, наиболее часта в Йемене (76%), Саудовской Аравии (64%) и Катаре (58%), также эта группа является обычной для Негевских бедуинов (62%). Среди других арабов она также очень распространена, особенно живущих в Леванте, т.е. палестинцев (38,4%), в Сирии (30%) и Ливане (25%). Среди еврейского населения J1c3dсоставляет 30% йеменских евреев, 20,0% ашкенази и 12% сефардов.
Предположительно, этому роду принадлежал легендарный Авраам.
Дагестан
Гаплогруппа J1 наиболее часта в Дагестане среди аварцев (58%), кубачи (99%), лезгинов (45%), даргинцев (69%),табасаранов (49%), кайтаки (85%), чеченцы (20%)
Северная Африка
В Северной Африке (на основании характерности YCAIIa22-YCAIIb22 среди алжирцев 35,0% и тунисцев 31%), J1 впервые попала в Эфиопию с распространением семитоязычных племён (Эритрея 11%, Эфиопия 9%, Эфиопия-Амхара (33,3%). J1 также с высокой частотой регистрируется в северных частях Судана (J-12f2(xJ2-M172): суданские арабы 45%, нубийцы 41%, копты39%, беджа 36%), и с меньшей частотой присутствует в районе Darfur (J-12f2(xJ2-M172): Masalit 6%, Fur 6%). Гаплогруппа J1может быть найдена по крайней мере у 20% египетских мужчин.
Европа
В целом частота J1 очень низка. Однако высокая плотность была зафиксирована в центральных Адриатических районах ИталииГоргано (англ.) (17,2%), Пескара (15%), в средиземноморской Паоле (11,1%), южносицилийской Рагузе (10,7%), на Крите (8,3%),Мальте (7,8%), Кипре (6,2%), в Греции (5,3%).
Гаплогруппа J2




Гаплогруппа J2 зародилась примерно 18000 лет назад на Ближнем
Востоке или Анатолии и затем распространилась на Балканы и во всем
Средиземноморье. Представители гаплогруппы J2 изобрели земледелие и
первыми стали одомашнивать животных, тем самым подарив миру
скотоводство. В центральную Европу некоторые представители гаплогруппы
J2 могли прибыть с одной из нескольких миграционных волн из Анатолии,
частично с греками, финикийцами, или с римскими легионерами и
поселенцами в начале нашей эры.
В Европе гаплогруппа J2 наиболее
распространена в Греции и Италии, где на eё долю приходится до 30 %, а
также в Турции и на Кавказе. На северном Кавказе в высокой концентрации у
ингушей (до 87%) и чеченцев (до 67%). В Индии с достаточно высоким
процентом (в среднем до 10%) встречается подгруппа J2b (snp M241).
Другая подгруппа J2a ограничена, главным образом, северо-западом
субконтинента. Вероятно часть из них попали туда с неолитическими
миграциями, а часть при индо-европейском вторжении.
Большое разнообразие J2 в турецком и южных европейских поселениях
объясняется исторической связью с Эгейской культурой и с Ближним
Востоком. Многие члены гаплогруппы J2 были ограничены прибрежными
средиземноморскими областями. Именно морская торговля вокруг
Средиземноморья, возможно, наравне с неолитической сельскохозяйственной
миграцией, способствовала распространению гаплогруппы J2 всюду в
средиземноморском мире...
Гаплогруппа C3

Генетический маркер "SNP M217", определяющий эту гаплогруппу возник 20
000 лет назад у человека который жил где-то в древних Восточноазиатских
поселениях. Он дал начало большому роду, который со временем распался
на племена и начал расселяться с территории современной Монголии. Это
расселение способствовало обособлению отдельных ветвей рода, среди
которых можно выделить две:
1) C3c (M48) – монгольские народы, тунгусо-маньчжурские народы, коренные
народы Восточной Сибири, тюркские народы Сибири и Центральной Азии. К
этому же роду относятся два таинственных изолированных народа – юкагиры и
айны, происхождение языка которых до сих пор остается спорным среди
учёных.
2) C3d (M407) – часть якутов и небольшая часть китайцев.
Генеалогии полагают, что это распространение во многом обязано
завоевательным походам Чингисхана в XII-XIII веках. В данной гаплогруппе
выделяется очень многочисленная и молодая ветвь, чей предок жил в эпоху
Чингисхана. Приблизительно до 16 миллионов мужчин по некоторым
подсчетам, живущих в Центральной и Восточной Азии принадлежит этой
ветви. Профессор Оксфордской академии Брайан Сайкс объявил
этот кластер родом Чингисхана, однако по данным российских
исследователей, это не совсем так, скорее всего, родоначальником данного
субклада является предок Чингисхана, возможно его прапрадед.
Гаплогруппа О3

Эта гаплогруппа возникла около 15 000 лет назад от мужчины, который,
вероятно, родился на территории современного Китая. Более 80% китайцев
являются потомками этого человека по мужской линии.
Взрывной популяционный рост гаплогруппы О3 связан с неолитической
революцией в Восточной Азии - выращиванием риса. Имея секрет технологии
рисоводства позволявшего прокормить большую численность населения, гаплогруппа О3 сумела
подавить окружающие популяции и вырвалась в родовые лидеры восточной
Азии. Этот демографический рывок произошел примерно в то же самое время,
что и аграрная зерновая революция в районе Плодородного Полумесяца на
Ближнем Востоке. При археологических раскопках в северном Китае было
обнаружено просо, начало распространения которого связывают с
междуречьем Тигра и Евфрата.
4000 лет назад рисоводство вместе с представителями гаплогруппы О3
достигло островов Индонезии Борнео и Суматры, и сегодня носители O3
обнаруживаются вплоть до отдаленной Полинезии.
Деятели генеалогии принадлежащие гаплогруппе O3
 Руководитель ДНК проекта O3 Денис Григорьев |  Автор проекта "Казахское Шежере" Аллиола Халидуллин |
Гаплогруппа Q

Гаплогруппа Q происходит от человека, родившегося в Сибири. Его
потомки Q1a-M3 были первыми исследователями Американского континента.
Несмотря на холодные температуры, часть гаплогруппы Q постепенно
продвигалась через свободную ото льда тундру с юга Сибири на
Северо-восток. Приблизительно 15 000 лет назад, они, дойдя до Чукотки,
сумели перебраться на соседний континент. Как им это удалось до сих пор
остается спорным вопросом. По одной из версий огромные массы воды в ту
пору были аккумулированы в леднике, и уровень мирового океана был на 100
метров ниже, чем сейчас. Следовательно, Сибирь и Аляска были соединены
перешейком, через который гаплогруппа Q благополучно перебралась в
Америку и продолжила свой путь на юг, создавая по пути великие
американские цивилизации Инков и Ацтеков.

Род Ашина, возможно происходил из ветви оставшейся в Сибири части
гаплогруппы Q, и сумел объединить под своим началом разнородные племена
(R1a-Z93, N1), положил начало созданию Тюркского каганата. По некоторым
данным, тюркский язык мог и быть языком рода Ашина, который восприняли
все объединенные в Каганате рода. После распада Тюркского каганата
правящая верхушка перебралась в отдаленную провинцию - в Хазарский
каганат, где и правила до 10 века. ДНК-генеалоги получили гаплотип
предполагаемых потомков рода Ашина - оказалось Q1b, и сейчас потомки
рода имеют свой ДНК - проект. Среди участников данного ДНК-генеалогического проекта имеются некоторые известные москвичи.
Гаплогруппы: описание древних родов и связь с археологическими культурами
Важная информация
Информация о происхождении и этнической принадлежности гаплогрупп на этом сайте не должна рассматриваться как неопровержимый факт, но, как это часто бывает в науке, в качестве модели в процессе постоянной эволюции на основе современных знаний и понимания (авторов). Всякий раз, когда прогресс генетики не может предоставить неопровержимые ответы, мы попытались предоставить наиболее вероятные и логические гипотезы на основе археологических, исторических и лингвистических доказательств. Эта страница постоянно обновляется, чтобы идти в ногу с недавними исследованиями и дает дополнительное понимание и исправление возможно ошибочных теорий.Введение в генетическую ДНК-генеалогию
Исследования ДНК позволили классифицировать всех людей на Земле в генеалогических группах, ведущих свое происхождение от одного общего предка с доисторических времен. Они называются гаплогруппы. Есть два вида гаплогрупп: отечески унаследованные по Y-хромосоме ДНК - Y-гаплогруппы, и наследуемые по материнской линии митохондриальной ДНК- мт-ДНК- гаплогруппы. Они указывают соответственно на происхождение по прямой мужской линии и прямой материнской линии.Y-ДНК гаплогруппы позволяют определить, являются ли два человека с одинаковой фамилией родственниками и действительно ли происходят от общего предка в недалеком прошлом (от 3 до 20 поколений). Это достигается путем сравнения гаплотипов через STR маркеры. Глубокое тестирование SNP позволяет вернуться гораздо глубже во времени, и определить древние этнические группы, к которым принадлежали предки по прямой мужской и прямой женской линии (например, кельтские, германские, финские, славянские, угорские, балтские, греко-римские, баскские, иберийские, финикийские, еврейские и др. рода).
В Европе гаплогруппы мт-ДНК довольно равномерно распределены по всему континенту, и поэтому не всегда могут быть связаны с легкостью с древними этносами. Тем не менее, иногда они могут выявить некоторые потенциальные заболевания, связанные с мутациями мт-ДНК. Некоторые мт-ДНК субклады связанны с еврейским происхождением, в частности, K1a1b1a, K1a9, K2a2a и N1b.
ДНК факты
Нуклеотиды являются алфавитом ДНК. Их всего четыре: аденин (А), тимин (Т), гуанин (G) и цитозин (C). Они всегда идут парами - А с Т, а G с C. Такие пары называются "пары оснований".46 хромосом ДНК человека состоит в общей сложности из 3000 миллионов пар оснований.
Y-хромосома содержит 59 млн. пар оснований, против 153 млн. на Х-хромосоме.
Митохондриальная ДНК находится вне ядра клетки и, следовательно, вне хромосом. Она состоит только из 16 569 пар оснований.
SNP (полиморфизм одного нуклеотида) является мутацией в одной паре оснований. В настоящее время лишь несколько сотен SNP определяют все человеческие гаплогруппы мт-ДНК и несколько тысяч для Y-ДНК гаплогрупп.
Y-ДНК гаплогруппы в генетической ДНК-генеалогии
Хронологическое и географическое возникновение Y-ДНК гаплогрупп
F => 55000 лет назад (на Ближнем Востоке)K => 45000 лет назад (в Южной Азии или Средней Азии)
KxLT (MNOPS) => 42000 лет назад (в Юго-Восточной Азии)
IJ => 41000 лет назад (на Ближнем Востоке)
NO => 33000 лет назад (в Юго-Восточной Азии)
P => 33000 лет назад (в Южной Азии)
T => 35000 лет назад (около Красного моря и в регионе Персидского залива)
J => 31000 лет назад (на Ближнем Востоке или в Анатолии)
R => 28000 лет назад (в Сибири или Южной Азии)
Q => 31000 лет назад (в Сибири)
E1b1b => 26000 лет назад (в Восточной Африке)
I => 25000 лет назад (на Балканах)
R1a1 => 16000 лет назад (в Сибири, на Урале или Центральной Азии)
R1b => 19000 лет назад (в Центральной Азии или в Закавказье)
J1 => 20000 лет назад (горы Загрос)
J2 => 25000 лет назад (в северной Месопотамии)
E-M78 => 18000 лет назад (на Африканском роге)
G => 26000 лет назад (в восточной части Ближнего Востока или Памире)
I2 => 22000 лет назад (на Балканах)
E-V13 => 10000 лет назад (на юге Ближнего Востока или в Северной Африке)
I2b => 13000 лет назад (в Центральной Европе)
N1c1 => 12000 лет назад (в Сибири или Восточной Азии)
I2a => 11000 лет назад (на Балканах)
G2a => 16000 лет назад (в Леванте или Анатолии)
R1b1a2 => 8000 лет назад (в районе Черного моря)
E-M81 => 9500 лет назад (в северо-западной Африки)
I2b1 => 9000 лет назад (в Германии)
I2a1 => 8000 лет назад (на юго-западе Европы)
I2a2 => 7500 лет назад (в Юго-Восточной или Восточной Европе)
I1 => 5000 лет назад (в Фенноскандии или Ютландии)
R1b-L21 => 4700 лет назад (в Центральной Европе)
R1b-S28 => 4600 лет назад (примерно в Альпах)
R1b-S21 => 5000 лет назад (в Центральной Европе)
I2b1a => меньше, чем 3000 лет назад (в Великобритании)
Европейские гаплогруппы
Гаплогруппа R1b

Гаплогруппа R1b является самой распространенной гаплогруппой в Западной Европе, достигая более 80% среди населения в Ирландии, Шотландии, Уэльс-западный, в Атлантической Франции и в Стране Басков. Также распространена в Анатолии и по всему Кавказу, в регионах России и в странах Центральной и Южной Азии. Кроме того, Атлантического и Северного побережья Европы, максимумы в долине рек на севере центральной Италии (более 70%), у осетин-дигорцев на Северном Кавказе (около 23%), в Армении (27%), у башкир на Урале (43%), Туркменистан (30%), хазарейцы народа Афганистана (25%), уйгуры Северо-Западного Китая (до 20%). R1b-V88, субклад широко распространенный в экваториальной Африке, встречается до 95% у мужчин в северном Камеруне.
У гаплогруппы R1b Анатолийское происхождение?
Происхождение гаплогруппы R1b не совсем ясно и по сей день. Некоторые из самых старых форм R1b находятся на Ближнем Востоке и в Закавказье. Гаплогруппа R1* и R2*, возможно, возникла в Сибири, на юге Центральной Азии (между Каспийским морем и Гиндукушем), либо в Южной Азии. Субклад R1 развился в R1b*, потом в R1b1* в северной части Ближнего Востока во время ледникового периода. Это произошло предположительно при переходе в северную Анатолию и на Кавказ в начале неолита, где появился субклад R1b1a. Ближневосточные остатки превратились в R1b1a (M18), который теперь встречается в низких частотах у ливанцев и у друзов. Финикийцы (которые пришли из современного Ливана) распространили эту гаплогруппу R1b1a и R1b1* в своих колониях, в частности, на Сардинии и в Магрибе.Субклады R1b1a1 и R1b1a2 (наиболее распространенные в Европе), тесно связаны с распространением индоевропейских языков, о чем свидетельствует их присутствие во всех регионах мира, где индоевропейские языки были в древности, от побережья Атлантики в Европе до Иранского нагорья, в том числе почти везде в Европе (за исключением Финляндии, Боснии и Герцеговины), Анатолии, Армении, Южной Сибири, во многих местах в Центральной Азии (в частности, СУАР, Туркменистан, Таджикистан и Афганистан), а также в Иране, Пакистане. История гаплогрупп R1b и R1a на начальном этапе были неразрывно связаны друг с другом. В то время как R1b1 найдено в таком месте, как Левант и Камерун, R1b1b скорее всего возник в восточной Анатолии.
Северный Кавказ и понтийско-Каспийские степи: индоевропейские связи
Современные лингвисты поместили прото-индоевропейскую прародину в Понтийско-Каспийские степи, различные географические и археологические области от устья Дуная до Уральских гор на востоке и на Северном Кавказе, на юге. Неолит, энеолит и ранний бронзовый века в культуре Понтийско-Каспийской степи были названы Курганной культурой (7000-2200 до н.э.) согласно гипотезе Марии Гимбутас, благодаря практике захоронения умерших под курганами ("курган") между последовательностью культур в этом регионе. Лошади были впервые одомашненные около 4000 до н.э. в степи, возможно, где-то в районе Дона и Нижней Волги, и вскоре стали определяющим элементом степной культуры. Во времена бронзового века- периода, известного как горизонт Ямной Культуры (3300-2500 до н.э.), крупного рогатого скота и овец, пастухи использовали вагоны для транспортировки пищи и палаток, которые позволяли им двигаться дальше в степь, порождая новый мобильный образ жизни, что в конечном итоге приведет к большим индоевропейским миграциям.Понтийско-Каспийские культуры степей можно разделить на западную часть, начиная от Дона до Днестра (а позже Дуная), и восточную, в Волго-Уральском регионе. Понтийские степи, вероятно, населенные мужчинами смешанных R1a и R1b линий, с более высокой плотностью R1b к северу от Кавказа, и R1a в северной степи и лесостепи.
R1b почти наверняка перешли из Северной Анатолии понтийско-Каспийской степи. Пока до конца не ясно, произошло это до, во время или после неолита. Регулярный поток R1b на Кавказе не может быть исключен. Генетическое разнообразие R1b больше всего в Закавказье, трудно отрицать, что R1b поселились и развивались там перед входом в степи мира. Означает ли это, что индоевропейские языки возникли в степи с людьми, R1a и R1b, что иммигранты слились в установленной культуре? Или что прото-индо-европейский язык появляется в северной Анатолии или в Закавказье, а уже затем распространился в степи с R1b? Или же еще прото-индо-европейцы впервые появляются в степи, как гибридный язык Кавказских / анатолийских R1b и R1a? Этот вопрос не имеет очевидного ответа, но, основываясь на древность и архаичность анатолийских языков(хеттский, палайский, лувийский, лидийский, и так далее), гипотеза происхождения прото-индо-европейского языка в Анатолии заслуживает доверия. Кроме того, есть документальные подтверждения слов, заимствованных из кавказских языков в индо-европейских языках. Это намного более вероятно, произошло, если бы прото-индо-европейский развился около Кавказа, чем в далекой степи. За R1b, следовательно, было распространение прото-индоевропейских языков в степи, а оттуда в Европу, Центральную Азию и Юго-Восточной Азии.
Майкопская культура и гаплогруппа R1b ведут в степи?
Майкопская культура (3700-2500 до н.э.) на Северном Кавказе, была культурным расширением Ямной культуры. Несмотря на это, как правило, она не считается частью понтийско-каспийской степной культуры благодаря своей географии, но на Северном Кавказе была тесно связана со степью, о чем свидетельствуют многочисленные керамика, золото, медь и бронза. Оружие и ювелирные изделия в современных культурах: Михайловка, Средний Стог Оба и Кеми. Связь между Северным Причерноморьем и Северным Кавказом старше Майкопского периода. Его предшественник, следующая культура (4400-3700 до н.э.), уже была в Суворово-Новоданиловке и в начале культуры Средний Стог, и даже старше аналогичной культуры Хвалынск на Волге. Это может быть период, когда R1b начали взаимодействие и смешение с R1a населением степей.Люди Ямной и Майкопской культуры использовали курганные захоронения с их умершими в положении лежа с поднятыми коленями и ориентированными по северо-восточной/юго-западной оси. Могилы были посыпаны красной охрой на полу а жертвенное животное хоронили рядом с людьми. Они также знали верховую езду, телеги, крупный рогатый скот и овец, использовали в их экономике, основанной на использовании медного / бронзового боевых топоров (в качестве молотов) и кинжалов. В самом деле, старые телеги и бронзовые артефакты находятся на Северном Кавказе, а уже оттуда распространялись по степи.
Майкоп был передовой культурой эпохи бронзы, на самом деле это одна из первых археологических культур, принявших участие в развитии металлообработки, и, следовательно, оружия из металла. Старинный меч в мире был найден в конце Майкопской культуры в залежах кургана 31. Его стиль напоминает длинный кельтский меч. Кости лошадей и изображения лошадей появляются уже в начале Майкопских могильников, предполагая, что в культуре Майкопа, возможно, сформировался степной тип людей, которые имели тесные связи с ними. Тем не менее, присутствие элементов культуры коренным образом отличается от степной культуры и в некоторых местах может означать, что Майкоп был гибридным по населению. Без анализа ДНК нельзя говорить, были ли эти две группы населения анатолийской R1b и кавказской G2a, или носители гаплогруппы R1a, которые поселились здесь позже. Две или три народности, возможно, даже сожительствовали бок о бок в различных населенных пунктах. Типичная кавказская Y-ДНК линия (например, G2a) не последовала примеру индоевропейских миграций, поэтому смешанные браки должны были быть ограничены, по крайней мере, ограниченны для индо-европейских мужчин с кавказскими женщинами, а не наоборот.
Майкопские люди являются теми, кто ввел внедрение примитивных колесных транспортных средств (телег) от Месопотамии до степей. Это был революционный способ жизни в степи, а в дальнейшем привел к развитию (конных) боевых колесниц около 2000 года до нашей эры. Кавалерия и колесницы играют важную роль в последующих индоевропейских миграциях, что позволяет им быстро и легко победить любого с кем они столкнулись. В сочетании с современным оружием и бронзовыми технологиями, западная ветвь (R1b) индо-европейцев от берегов Черного моря являются превосходными кандидатами на то, что это таинственный народ моря, который ворвался к восточному берегу Средиземного моря в период второго тысячелетия до нашей эры.
Распространение индо-европейского языка хеттов в центральной Анатолии произошло спустя несколько веков после исчезновения Майкопской культуры. Обратный переход от Северного Кавказа до северной Анатолии, очень вероятно, произошел в этот периоде экспансии. Не вызывает сомнений то, что хетты использовали колесницы, изобретенные в Волго-Уральской степи. R1a были обнаружены с низкой частотой в Армении и Северной Анатолии, что дает основания предположить, что гибридные группы R1a, R1b из Волго-Уральского региона мигрировали в этот регион в промежутке между 2000 г до н.э. и 1650 до н.э. Майкопскую и Ямную культуры сменила Срубная культура (1600-1200 до н.э.), возможно, представленная из носителей гаплогруппы R1a1 северных и восточных степей распространившихся к берегам Черного моря.
Европейские субклады гаплогруппы R1b
Наличие у Индоевропейцев бронзового оружия и лошадей, дали им огромное преимущество перед коренными обитателями Европы, а именно перед гаплогруппой I (потомки первых европейских кроманьонцев), и первых неолитических культур пастухов и фермеров (G2a, J2, E-V13 и T). Это позволило гаплогруппам R1a и R1b заменить большинство старых мужских линий в Западной Европе, хотя женские линии, кажется, были менее затронуты.По сравнению с индо-иранским вторжением в Южную Азию показательно, что около 40% мужских линий северной Индии принадлежат гаплогруппе R1a, но менее 10% женских линий может быть названо индо-европейскими по происхождению. Влияние индоевропейцев было более тяжелым в Европе, потому что европейское общество 4000 лет назад было менее развито с точки зрения сельского хозяйства, технологии (без бронзового оружия) и плотность населения ниже, чем в цивилизациях долины Инда. Это особенно справедливо в западноевропейских странах, где сельское хозяйство пришло намного позже, чем на Балканы и в Центральную Европу. Греция, в целом Балканы и Карпаты- были самые передовые европейские страны в то время и меньше всего пострадали в плане замены гаплогрупп. Местные европейские Y-ДНК гаплогруппы (I1, I2a, I2b) также сохранились лучше в тех регионах, которые были более труднодоступны и менее гостеприимны - как в Скандинавии, Британии, Сардинии или в Динарских Альпах.
Первые нападения степных людей на Балканы произошли между 4200 до н.э. и 3900 до н.э., когда наездники перешли Днестр и Дунай и, видимо, разрушили города Варну и Караново культуры VI в Восточной Румынии и Болгарии. Изменение климата в результате холодных зим в течение этого периода, вероятно, подтолкнуло степных пастухов искать мягкие пастбища для своего скота, что привело к голоду и внутренним беспорядкам в дунайской и балканской общинах. Последовавшая за этим Чернаводова культура (4000-3200 до н.э.) и Эзеро культура (3300-2700 до н.э.), кажется, были из смешанного населения степных иммигрантов и людей из старых поселений. Эти степные иммигранты, вероятно имели смесь обоих гаплогрупп R1a1 и R1b. Многие дунайские фермеры также мигрировали в Кукутень-Триполье в Восточных Карпатах, в результате чего население направилось на северо-восток расширяясь до долины Днепра, в результате чего Y-гаплогруппы E-V13, J2b и T есть в настоящее время в центральной Украине. Это преждевременное движение индоевропейских народов на запад было довольно ограниченным, в связи с отсутствием бронзового оружия и организованной армии в то время, и было действительно возможно только благодаря климатическим катастрофам. Карпатская, дунайская и балканская культуры были слишком густонаселенными и технологически продвинутыми, чтобы обеспечить массовую миграцию.
Бронзовый век анонсирует новое развитие. Носители гаплогруппы R1a1, вероятно, были первыми, кто успешно проникнул в самое сердце Европы, в составе культуры шнуровой керамики (3200-1800 до н.э.) как естественное расширение на запад Ямной культуры. Они пошли на запад, в Германию и Скандинавию. Анализ ДНК из продовольственной культуры Иелау подтверждает наличие гаплогруппы R1a1 в центральной Германии около 2600 до нашей эры. Эта культура вполне могла бы расширится в лесостепи, или в северной окраине Ямной культуры, где R1a линии были распространены.
Гаплогруппа R1b1a2, как полагают, прибыли в Центральную и Западную Европу около 3000 г. до н.э., идя вверх по Дунаю от побережья Черного моря, другой поток через Анатолию и Кавказ и третий, возможно, через северную Африку на Пиренеи. Археологические и генетические данные (распределение субклада R1b), указывают на несколько последовательных волн в направлении Дуная между 2800 до н.э. и 2300 до н.э. (начало Унитецкой культуры). Интересно отметить, что это также соответствует концу культуры Майкопа (2500 до н.э.) и Обской культуры (2200 г. до н.э.) на северном побережье Черного моря, замена культур происходит из миграций со стороны северных степей. Поэтому может быть предусмотрено, что (в основном) R1b население из северной части Черного моря мигрировали на запад из-за давления со стороны других индоевропейских народов (R1a) с севера, в виде растущей прото-индо-иранской ветви, связанной с современными Абашевской и Полтавской культурами.
Возможно, что культура Колоколовидных Кубков (2800-1900 до н.э.) была уже индоевропейской (хотя они были под влиянием культуры шнуровой керамики), потому что они были в непосредственной связи с родной мегалитической культурой. Есть вероятность, что кубки и лошади обнаруженные в Западной Европе в этот период были результатом торговли с соседними индоевропейскими культурами, в том числе первой волны R1b в Центральной Европе. Тем не менее, нельзя отрицать, что следующая Унитецкая культура (2300-1600 до н.э.), культура Курганов (1600-1200 до н.э.), Культура полей погребальных урн (1300-1200 до н.э.) и Гальштатская (1200-750) культуры были связаны с распространением R1b в Европе, они резко внедряют новые технологии и внедряют совершенно иной образ жизни.
Действительно ли индо-европейцы вторглись в Западную Европу?
Сторонники модели преемственности палеолита или неолита утверждают, что бронзовые технологии и лошадей можно было бы импортировать у западных европейцев от их восточноевропейских соседей, и что фактически индоевропейского вторжения могло не быть. Труднее понять, как Италийские, кельтские и германские языки были приняты Западной и Северной Европой без хотя бы небольшого вторжения. Было высказано предположение, что индоевропейская (IE) ветвь языков просто распространяется через контакт, как и технологии, или потому, что он был языком небольшой элиты и, следовательно, его принятие воспринимается как престижное. Однако людям не просто изменить язык, потому что это звучит лучше, или более престижно. Даже в наши дни, когда есть учебники, словари, обязательные языковые курсы в школе, курсы в частных языковых школах для взрослых и многоязычные телевизионные программы, большинство людей не может свободно овладеть иностранным языком, принадлежащим к другой языковой семье или группе. Языковой разрыв между пред-индо-европейскими наречиями был примерно таким же большим, как между современным английским и китайским языками. Английский, греческий, русский и хинди являются индо-европейскими языками и, следовательно, легче учатся в общей динамике, чем в не-ИЕ, как китайском, арабском или венгерском. С лингвистической точки зрения, только широкомасштабной миграцией ораторов можно объяснить массовое принятие индо-европейских языков в Западной Европе - оставляя только басков как пережиток языков неолита.Одним из важных археологических аргументов в пользу замены неолитических культур индо-европейской культурой в эпоху бронзового века исходит из керамических стилей. Внезапное появление бронзовой технологии в Западной Европе совпадает с керамикой и неожиданно становится все более простым и менее украшенным, как и в понтийских степях. До тех пор, гончарные изделия были постоянно в эволюционировании в сторону большей сложности и деталей более чем 3000 лет. Люди не просто решили вернуться к более примитивным стилям. Возможно, одно изолированное племя могло экспериментировать с чем-нибудь попроще, но каковы шансы, что далекие культуры Иберии, Галлии, Италии и Великобритании все решили провести такой невероятный сдвиг примерно в то же время? Лучшее объяснение, что этот новый стиль был введен иностранными захватчиками. В данном случае это не просто спекуляции, есть достаточно доказательств, что это простая керамика характерная связана с появлением прото-индо-европейского языка.
Кроме керамики, археология дает достаточно доказательств, что ранний бронзовый век в Центральной и Западной Европе совпадает с радикальным сдвигом в производстве продуктов питания. Сельское хозяйство испытывает резкое сокращение в обмен на повышенное внимание в одомашнивании скота. Это также период, когда лошади все чаще используются и коровье молоко потребляется регулярно. Даже после введения сельского хозяйства около 5200 до н.э., Буг, Днестр - культуры и поздние культуры степей характеризуются в экономике доминированием животноводства, только с ограниченным сельским хозяйством. Эта модель расширяет свою деятельность в Европе, именно в то же время, как бронза появляется.
Религиозные верования и искусство прошли полный переворот в бронзовом веке в Европе. Неолитические общества на Ближнем Востоке и в Европе всегда поклонялись женским фигуркам в форме культа плодородия. Степные культуры, наоборот, не производили женских фигурок. С распространением бронзовых технологий в долине Дуная в Западной Европе, символы плодородия и плодовитости постепенно исчезают и заменяются скульптурами домашних животных.
Другим признаком того, что индо-европейские народы пришли в большом количестве в регион Центральной и Западной Европы можно найти в практике захоронений. Европейцы неолита или кремировали своих умерших (например, Кукутень-трипольская культура) или хоронили их в коллективных могилах (это было в случае мегалитических культур). В степи, каждый человек был похоронен отдельно, и высокопоставленные могилы помещали в погребальную камеру, увенчанную круговой насыпью. Обычно кладется оружие (булавы, топоры, кинжалы), лошади костей, и демонтированные телеги (или более поздние колесницы). Эти характерные курганы известны как курганы в понтийских степях. Мужчины получили больше роскошных гробниц, чем женщины, даже среди детей, и различия в иерархии очевидны между захоронениями. Индоевропейцы были сильно иерархическим мужским обществом, в отличие от более равноправного и матрилинейного общества культур Старой Европы. Распространение статуса доминирующих мужчин в Центральной Европе в эпоху бронзы является явным признаком, что правящая элита теперь стала индоевропейской. Эта практика также распространяться в Центральной Азии и Южной Сибири, двух регионах, где R1a и R1b линий находятся в настоящее время, как в Центральной Европе. Церемония захоронения одного из самых эмоциональных и личных аспектов культуры. Весьма сомнительно, что люди будут менять своих предков практикой "просто сделать, как у соседей". На самом деле, различные погребальные методы сосуществовали бок о бок во время европейского неолита и энеолита. Принятие некоторых элементов иностранной культуры как правило, происходит, если одна цивилизация показывает для соседних культур свое превосходство. Этот процесс называется "аккультураций". Однако ничто не указывает, что степная культура была настолько культурно превосходной, чтобы мотивировать весь континент, даже Атлантические культуры на протяжении 2000 км от понтийских степей, к оставлению многих таких фундаментальных символов собственной древней культуры, и даже свой собственный язык. На самом деле, старая Европа была гораздо более изысканной в своей керамике и ювелирных изделий, чем "грубые" люди степи. У Индо-европейского первенства были культурные преимущества, а также военные, благодаря лошадям, бронзовое оружие и нормы профессиональной оценки отдельных героических подвигов во время войны (эти этические ценности, как известно из старых текстов, таких как Ригведа, Авеста, или микенской и Хеттской литературы).
После лингвистики и археологии, третья категория доказательств исходит от генетики. Впервые было предположено, что R1b - родом из Западной Европы, потому что это место, где она была наиболее распространенной. С тех пор было доказано, что гаплотипы гаплогруппы R1b отображаются выше микросотелитных разнообразий в Анатолии и на Ближнем Востоке, чем в Европе. Европейские субклады также более поздние, чем на Ближнем Востоке и в Центральной Азии. Основные европейские субклады, R-P312/S116 только восходят от 3500 до 3000 до нашей эры. Это не значит, что самые старые общие предки этой линии прибыли в Западную Европу в этот период, но это первый человек, который нес мутацию R-P312/S116 жил по крайней мере 5000 лет назад, предположительно где-то около Черного моря. В любом случае этот срок слишком поздний для происхождения из палеолита, мезолита или даже неолита для R1b в Европе. Открытие того, что считалось «Европейской линией» в Центральной Азии, Пакистане и Иране вбило "последний гвоздь в гроб" происхождения палеолитических R1b в Западной Европе, и подтвердило индоевропейские гипотезы.
Все элементы сходятся в пользу крупномасштабной миграции на основе верховой езды индоевропейских народов в Западной Европе между 2500 до 2100 до н.э., что способствует смене образа жизни неолита и энеолита - по сути новая культура бронзового века, с простой глиняной посудой, земледелием, скотоводство приобретает новые ритуалы и новые значения (для мужской линии общества, воинов и героев), которые не развиваются из местных предшественников.
Эти прото-итало-кельто-германские R1b люди поселились вокруг Альп 2300 году до н.э., и, судя по распространению бронзовых работ, достигли Иберии примерно в 2250 до н.э., Великобритании к 2100 году до нашей эры и Ирландии в 2000 году до нашей эры. Это первая волна R1b предположительно осуществлялась R1b-L21 линией в большем количестве, так как они встречаются повсеместно в западной, северной и центральной Европе. Второе расширение R1b состоялось с культурой полей погребальных урн / Гальштатской культуры около 1200 г. до н.э., направляясь на запад до Атлантического океана, к северу до Скандинавии, и на восток до Греции и Анатолии.
Новая культура бронзового века процветала вокруг Альп (от Унитецкой до начала Гальштатской) благодаря обилию металла в регионе, а также заложены основы для классической кельтской культуры.
Альпийские кельты Гальштатской культуры, связанные с S28 (ака U152) мутацией. Отделились и (в том числе S28/U152) вступили в Италию в 1200 г. до н.э., но было, конечно, несколько последовательных волн, о чем свидетельствуют более поздние прибытия Цизальпинских кельтов. Белги были также S28/U152, расширение культуры - Латенская на север, дальше за Рейн, Мозель, Мез и впадающие реки.
Одна из распространенных языковых черт в галльских / Бриттских и кельтских языках, связанных с расширением Гальштатской культуры указывает, что они поменяли отдельные звуки. Они, как известно лингвистам, считаются P-кельтской ветвью. Считается, что это изменение произошло в связи с неспособностью произносить звук, естественный для индоевропейского населения Центральной Европы, Галлии и Италии, которые были носителями афро-азиатских диалектов, которые произошли от ближневосточных языков. Этруски, хоть и более поздние пришельцы из Леванта, также вписываются в эту категорию. Недавно была высказано гипотеза, что кельтские языки заимствовали часть своей грамматики в афро-азиатских языках. Это изменение могло произойти, когда прото-итало-кельты перешли из степи в бассейне Дуная и смешались с населением ближневосточных фермеров принадлежащих гаплогруппам E-V13, T, G2a и J2b. Однако такой ранний переход не может объяснить, почему слова кельтского языка развились в Ирландии и Иберии. Более вероятно, что смешение произошло после того, как итало-кельты впервые распространились во всех странах Западной Европы. R1b-S28/U152 в связи с P-кельтским означает, что произошел переход вокруг Альп и Италии после 1200 г. до нашей эры.
R1b-S21 (ака U106) найден в высоких концентрациях в Нидерландах и Северной Германии. Его присутствие в других странах Европы могут быть отнесены к 5-й и 6-м векам германских миграций. Фризы и саксы распространили эту гаплогруппу на Британских островах, франки в Бельгии и Франции, и лангобарды в Австрии и Северной Италии. Высокая концентрация S21/U106 вокруг Австрии могла возникнуть там, в Гальштатский период, или возникла вокруг Черного моря и перешла туда в Гальштатский период. На самом деле, на юге Германии и Австрии, вместе взятые имеют самое высокое разнообразие R1b в Европе. Кроме S21, три важнейших первых субклада уровня R1b1b2a1b (L21, S28, M167) находятся в этом районе в разумных частотах чтобы усмотреть распространение из Унитецкой в Гальштатскую культуру с остальной Западной Европой.
Как R1b пришли на смену большинству старых линий в Западной Европе?
До недавнего времени считалось, что R1b зародилась в Западной Европе из-за своего максимального присутствия в регионе и по сей день. Считалось, что R1b - представители палеолита Европы (европейские кроманьонцы), которые нашли убежище в франко-Кантабрийском регионе на пике последнего ледникового периода, то реколонизация Центральной и Северной Европы после отступления ледяного покрова. Филогенией R1b доказано, что этот сценарий не существовал, потому что старшие клады R1b последовательны в Центральной Азии и на Ближнем Востоке, а более молодые в Западной и Северной Европе. Существовал четкий градиент с востока на запад отслеживания миграции R1b. В этом возрасте основной переход от берегов Черного моря до Центральной Европы также произошел в соответствии со сроками индоевропейского вторжения в Европу, которые совпадают с появлением культур бронзового века в Западной Европе, и распространением итало-кельтских и германских языков.Историки и археологи долгое время утверждали была ли индоевропейская миграция массированным вторжением, или, скорее, культурная диффузия языка и технологии распространялись только на небольшое количество пришельцев. Ответ на этот вопрос вполне может быть "нет". Сторонники теории диффузии хотели бы заставить нас думать, что R1b является родным субкладом для Западной Европы, а только R1a представлять индоевропейцев. Проблема в том, что гаплогруппа R возникает в Сибири или Южной Азии, и R2 по-прежнему ограничено Центральной и Южной Азии, в то время как обнаруженные древние линии R1a в Европе, а старшие субклады из R1b - в Азии. Возраст субклада R1b в Европе совпадает с бронзовым веком. R1b, следовательно, должна заменить большинство местных Y-ДНК линий в Европе с бронзового века и позже.
Тем не менее, массовая миграция и почти полная аннигиляция населения палеолита вряд ли может быть предусмотрена. Западные европейцы есть в Ирландии, Голландии, Португалии или Аквитании, несмотря на все регионы, где R1b является доминирующим. Аутосомные исследования ДНК подтвердили, что западноевропейское население далеко не однородно. Много материнских линий (мтДНК) также, вероятно, происхождением из палеолита (например, H1, H3, U5 и V), основанные на древних ДНК. То, что многие люди забывают о том, что есть и не нуждается в крупномасштабных исходах для мужской линии заменить достаточно быстро. И вот почему.
Многоженство. В отличие от женщин, мужчины не ограничены в количестве детей, они могут производить потомство. Мужчины как правило, имеют больше детей. Это было справедливо в примитивных обществах, где полигамия часто была нормой для вождей и королей.
Статус и питание. Оснащенные бронзовым оружием и лошадьми, индоевропейцы легко покорили неолитических фермеров и потом с еще большей легкостью охотников-собирателей Европы. Если они не истребляли коренных европейских мужчин, новички стали бы новым правящим классом, с Множеством местных царей, вождей и знати (кельты и германцы жили в маленьких сельских общинах с вождем, отдельная часть небольшого племени во главе с королем) с более высокими репродуктивными возможностями, чем в среднем.
Вторжение армии как правило, имеют гораздо больше мужчин, чем женщин. Люди должны поэтому найти женщин в покоренном населении. Войны ведутся людьми, и проигравшие имеют более тяжелые потери, в результате чего все большее число женщин попадает в распоряжение победителей.
Индоевропейцы были воинственным народом с сильной героической смелостью, подчеркивая честь и воинские доблести. Их превосходная технология (металлическое оружие, транспортные средства и герои) и отношение к жизни позволило бы им победить любую группу населения, которая не имеет организованной армии с металлическим оружием (то есть никому, кроме ближневосточных цивилизаций).
Есть ли генетическая предрасположенность к зачатию мальчиков у R1b?
Основная роль Y-хромосомы в теле человека состоит в запуске развития организма по мужскому типу. Гаплогруппы определяются на основе дифференциации мутации Y-хромосомы в основном в негенных областях. Каждая мутация теоретически способна отрицательно повлиять на производство спермы и подвижность сперматозоидов, если она выпала на участок с геном, например SRY. Предварительные исследования уже установили связь между определенной мутацией в Y-гаплогруппе и увеличением или уменьшением подвижностью сперматозоидов, чаще всего эти мутации патологические и приводят мужскому бесплодию. Чем выше подвижность, тем выше шансы на зачатие мальчика. Абсолютно исключено, чтобы R1b или любая другая гаплогруппа древнее 5 000 лет, имела преимущество в сторону большего мужского потомства. Увеличение или уменьшение подвижности сперматозоидов вследствие мутаций скорее могут быть характерны для отдельных молодых групп в разных гаплогруппах и субкладах.В любом случае, если на начальном этапе в этносе по какой-либо причине будет рост носителей отдельной гаплогруппы, то скорее всего в последующем, в этой популяции сохраниться такая же пропорция - это называется эффектом основателя. Смещения существуют только по отношению к другим гаплогруппам и обнаружены в одной и той же популяции. Вполне возможно, что сравнительно недавно R1b субклад Западной Европы имели значительное преимущество по сравнению со старыми гаплогруппами в этом регионе, в частности гаплогруппами I2 и G2a.
Замена мужской линии по этой модели быстро становится экспоненциальной. Представьте себе 100 индо-европейских мужчин завоевывает 1000 коренных европейцев (соотношение 1:10). Военные потери привели к более высокой доли женщин в покоренном населении. Давайте предположим, что уцелевшее население состоит из 700 женщин и 300 мужчин. Давайте предположим, что победоносные индо-европейские мужчины в конечном итоге, имеют в два раза больше детей, достигнув зрелого возраста, чем мужчины побежденного племени. Существует ряд причин для этого. Победители займут больше жен, наложниц или даже изнасилуют женщин побежденного племени. Их высокий статус будет гарантировать их больше богатства и, следовательно, более качественное питание для своего потомства, увеличивая шансы на достижение совершеннолетия и продолжения своего рода. Потомства в соотношении 2 к 1 для мужчин на самом деле является консервативной оценкой, так как это полностью возможно, что в бронзовом веке чувства привели бы к уничтожению большинства мужчин на проигравшей стороне (как свидетельствует Ветхий Завет). Тем не менее, это займет всего несколько поколений на победу Y-ДНК линий, чтобы стать большинством. Например, если в первом поколении индоевропейцев было два оставшихся в живых сына на человека, в отношении только одного человека на коренных жителей, число индоевропейских отцовских линий будет проходить в 200 человек во втором поколении, 400 в третьем, 800 в четвертом и 1600 в пятом, и так далее. За это время коренным линиям грозит только стагнация на 300 человек для каждого поколения.
На основе такого сценария, R1b линии быстро подавили бы местные линии. Даже если у индоевропейских завоевателей было лишь немного больше детей, чем у местных мужчин, R1b линия станет доминирующей в течение нескольких веков. Кельтская культура продолжалась более 1000 лет в континентальной Европе до римского завоевания, чтобы положить конец привилегии от вождей и знати. Это более чем достаточно по времени для R1b линий достичь от 50 до 80% населения.
Современные частоты R1b формируют градиент по краю Атлантической Европы (высокий процент) в Центральной и Восточной Европе (самый низкий), вновь поднимается на анатолийской родине. Это почти наверняка связано с сельским хозяйством, которое лучше развито в Восточной и Центральной Европе, а затем, с более высокой плотностью населения, в результате чего R1b вторженцев здесь меньше, чем на Западе. Кроме того, другие индо-европейские культуры шнуровой керамики (R1a) уже прошли путь от современных России и Украины на запад, в Германию и Скандинавию. Было бы трудно для R1b людей, чтобы конкурировать со своими двоюродными братьями R1a, кто разделяет аналогичную технологию и культуру. Предварительно кельто-германские R1b поэтому вынуждены были уходить дальше на запад, прежде всего в Альпы, то есть обгоняя, те малонаселенные части Западной Европы.
Греко-анатолийские ветви гаплогруппы R1b
Хетты (2000-1200 до н.э.) были первыми индоевропейцами сделавшими вызов (и потерпевшими поражение) с могучей месопотамской и египетской империями. Хеттский правящий класс был доказано ответвлением от Майкопской культуры, который завоевал царство хаттов. Северная Анатолия, возможно, также была заселена индоевропейцами, которые позже основали Майкопскую культуру и распространили свой язык и культуру в Понтийско-Каспийской степи. Какой бы не был путь, северные анатолийцы бронзового века - это индоевропейские мигранты, и несомненно принадлежали по большей части к гаплогруппе R1b1a2. Хатты, возможно, были из некоторых старых ближневосточных R1b, смешавшихся с другими гаплогруппами, распространенными в Анатолии в настоящее время (G2a, J2a и E-M78).Легендарная Троя вполне могла быть индоевропейской колонией для обеспечения торговых путей между Черным морем и Эгейским. Троянцы были лувийским перевалочным пунктом, связанным с хеттами (отсюда индоевропейские топонимы), с доказанными культурными связями с культурой Понтийско-Каспийской степи. Первые упоминания о Трое восходят к 3000 г. до н.э., в самом развитии Майкопской культуры, и во время производства первых галер. Учитывая раннее основание Трои, скорее всего, из двух индоевропейских гаплогрупп это были R1b1a2, а не R1a1a1.
Греческие R1b входят во многие разновидности: R1b1 с Ближнего Востока, R1b1b из Анатолии, и Европейские R1b1b2, в том числе прото-кельтские S116/P312 и Гальштат-кельтские S28/U152. Наличие R1b1b2 в Греции может быть связано с вторжением Дориан, как полагают, произошло в 12 веке до нашей эры. Дорийцы возможно были связаны с троянцами и хеттами, принадлежащими к древнейшим индоевропейским языковым группам, или прото-кельтами из Центральной Европы и долины Дуная. Среди их Y-ДНК линий было преимущественно R1b1b или R1b1b2. Дорийцы могут быть потомками первых (R1b), степных кочевников, которые обосновались в восточной части Балканского региона (Чернавода и Эзеро культуры) и не продолжали миграции вверх по Дунаю в Центральную и Западную Европу.
Греческие и анатолийские R1b-S28 линии могут быть связаны с кельтским вторжения в 3 веке до н.э., но более вероятно, с римской оккупацией. Старые клады из R1b, такие как R1b1 или R1b1a лишь в незначительном количестве и пришли с J2 и E1b с Ближнего Востока. Микенцы могли бы принести R1b1b2 в Грецию, но их происхождение можно проследить до Сейсмино-Турбинского феномена культуры северной лесостепи, которые были в большей степени племена из N1c1 и R1a1.
Центрально-азиатские субклады R1b
На заре цивилизаций группы R1b людей, как полагают, мигрировали из региона Каспийского моря в Центральную Азию, где она превратилась в R1b1b1 (M73) субклад. Эта разновидность R1b встречается почти исключительно в очень специфических местах у населения Центральной Азии. Самый высокий процент обнаружен среди уйгуров Синьцзяна (до 20%) на северо-западе Китая, у хазарейцев народа Афганистана (до 30%) и у башкиров Абзелиловского района (до 43%) в России (граница с Казахстаном).В Центральной Азии R1b1b1 может соответствовать тохарской ветви индоевропейцев. Вполне возможно, что тохары отделились от основной линии R1b в начале 7000 г. до н.э. На протяжении столетий некоторые группы этих кочевых племен мигрировали около Южного Урала, другие возле бассейна реки Тарим (Синьцзян) и в южной части Центральной Азии. Другая теория состоит в том, что группа ранних наездников из культуры Репина (3700-3300 до н.э.) мигрировали из Дон-Поволжья в горы Алтая, основав культуру Афанасьево (ок. 3600-2400 до н.э.), а затем переместились на юг в Тарим.
Мумии светловолосых людей европеоидной расы были найдены в бассейне Тарим, самые старые из которых датируются 1800 до нашей эры. Современные жители бассейна реки Тарим, уйгуры, относятся тому же R1b-M73 субкладу (до 20%) и R1a1 (около 30%). Это может означать, что они сформировались из гибридных R1b и R1a обществ к тому времени и достигли бассейна реки Тарим. Но R1a1 могли бы также прибыть самостоятельно в течение более поздних миграций культур Андроновского круга (ок. 2000 до н.э.), или гораздо позже, через несколько кочевых скифо-иранских или тюркских племен (после 700 г. до н.э.)
Гаплогруппа R1a1

Гаплогруппа R1a, как полагают, была доминирующей гаплогруппой среди северных и восточных индоевропейских народов, которые частью перешли в индо-иранские (R1a1-Z93), частью в греческие из микенской, фракийской, балтийской и славянских ветвей. Прото-индо-европейцы возникли в Ямную культуру (3300-2500 до н.э.), в Понтийской-Каспийской степи между современной Украиной и юго-западом России. Их расширение связано с одомашниванием лошадей в степях Евразии, и изобретением колесницы (см. выше R1b).
В восточной части понтийско-прикаспийские степи в значительной степени связаны с индо-иранскими и балто-славянскими ветвями индо-европейских языков. На основании археологических, языковых и генетических данных, можно сказать, что скотоводы-кочевники, жившие в северных русских степях и лесостепях 7000 лет назад были преимущественно R1a по отцовским линиям.
В настоящее время, высокие частоты гаплогруппы R1a находятся в Польше (56% населения), Украине (от 50 до 65%), европейской части России (от 45 до 65%), Беларуси (45%), Словакии (40%), Латвии (40 %), Литве (38%), Чехии (34%), Венгрии (32%), Хорватии (29%), Норвегии (28%), Австрии (26%), Швеции (24%), северо-восток Германии (23%) и Румынии (22%).
Германские субклады гаплогруппы R1a
Первое расширение гаплогруппы R1a1 состоялось на запад с распространением шнуровой керамики (3200-1800 до н.э.) с родины Ямной культуры. Это была первая волна R1a1 в Европе, которая отвечает за наличие этой гаплогруппы в Скандинавии, Германии и часть R1a1 в Чешской Республике, Словакии, Венгрии и Польше. Высокая распространенностьгаплогруппы R1a1 в балто-славянских странах в настоящее время является не только за счет расширения шнуровой керамики, а также длинного ряда позднейших миграций из России, последняя из которых состоялась с 1 по 5-й века нашей эры.Германская ветвь индоевропейских языков, вероятно, произошла от слияния одной из культур гаплогруппы R1a1 (на прото-славянском языке), а позднее прибытии итало-кельтского субклада R1b из Центральной Европы. Это подтверждается тем фактом, что германский народ гибридный из доли гаплогруппы R1a1 и большей частигаплогруппы R1b1a2, что эти две гаплогруппы пришли по отдельным маршрутам в разное время, а также по лингвистике Прото-германских языков, который имеет сходство с италийскими, кельтскими и славянскими языками.Гаплогруппа R1a1 Культуры Шнуровой керамики смешались с германскими аборигенами I1 в бронзовом веке (1800-500 до н.э.). R1b предположительно достигли Скандинавии позже миграцией на север от современной Гальштатской культуры (1200-500 до н.э.). Первый подлинный германский язык, по оценкам лингвистов, возник около (или после) 500 г. до н. Это подтверждает, что он произошел от смеси Гальштатского прото-кельтского населения и праславян Культуры шнуровой керамики. Уникальность некоторых из германских слов указывает на заимствования из общего индо-европейского языка. Кельтский язык, как известно, имеет много заимствованных слов из афро-азиатских языков, от ближневосточных иммигрантов в Центральной Европе. Тот факт, что современная Скандинавия состоит из примерно 40% I1, 20% R1a и 40% R1b подтверждает идею о том, что германская этническая принадлежность и язык носит три-гибридный характер со времен железного века.
Балтийские субклады гаплогруппы R1a1
Ветвь Балтийского, как полагают, произошли от Фатьяновской культуры (3200-2300 до н.э.), северо-восточный ареал культуры шнуровой керамики. В ранний бронзовый век кочевники гаплогруппы R1a1 из северных степей и лесостепей смешиваются с коренным уральским населением (N1c1 линий) в этой области. Это подтверждает сильные позиции как R1a, так и N1c1 гаплогрупп из южной Финляндии, в Литве и прилегающей части России.Славянские субклады гаплогруппы R1a1
Происхождение славян по прямой мужской линии R1a восходит к приблизительно 3000 г. до нашей эры. Славянские ветви дифференцированы, когда сама культура шнуровой керамики (см. германские ветви выше) поглощается Кукутень-трипольской культурой (5200-2600 до н.э.) из Западной Украины и северо-востоке Румынии, которая, кажется, была представлена в основном из I2a2 линий происходящих непосредственно из палеолита европейцев, с небольшой примесью северо-африканских иммигрантов (в частности, E-V13 и T). Таким образом, появились гибридные культуры шаровидных амфор (3400-2800 до н.э.) на территории нынешней Украины, Белоруссии и Польши. Это, безусловно, в период I2a2, E-V13 и T с распространением (наряду с R1a) вокруг Польши, Беларуси и западной части России, и объясняет, почему восточные и северные славяне (и литовцы) имеют значительную частоту гаплогруппы I2a2 с небольшим количеством E1 и Т. Через несколько веков, эта гибридизованная культура исчезла в доминирующей культуре шнуровой керамики.Период культуры шнуровой керамики (1700-1200 до н.э.) сопровождался лужицкой культурой(1300-500 до н.э.), чернолесовой (1025-700 до н.э.) и Милоградской (600 до н.э.-100 г. н.э.) культур на северо-востоке. Последняя важная славянская миграция, как полагают, произошла в 6-ом веке нашей эры, из Украины в Польшу, Чешскую Республику и Словакию, заполняя вакуум, оставленный от восточных германских племен, которые вторглись в Римскую империю.
Исторически сложилось, что никакая другая часть Европы не была захвачена большее количество раз, чем степных народы на Балканах. Хронологически первые захватчики R1a пришли на запад с расширением культуры шнуровой керамики (от около 3200 до н.э.), потом вторжение микенцев (1600 до н.э.), а затем фракийцы (1500 до н.э.), иллирийцы (около 1200 до н.э.), Гунны и аланы (400 н.э.), авары, болгары и сербы (все около 600 н.э.), и мадьяры (900 н.э.). Эти народы возникли из разных народов евразийских степей, между Восточной Европой и Центральной Азией, что объясняет, почему такое высокие разнообразие STR находится в пределах Балканских R1a в настоящее время. Это пока не представляется возможным определить этническое происхождение для каждой разновидности R1a, кроме того, что они связаны с племенами из степей Евразии в какой-то момент в истории.
Индо-иранские ветви гаплогруппы R1a1
Прото-индо-иранские языки, люди, которые позже назвали себя «ариями» в Ригведе и Авесте, возникла в синташтинской-Петровка культуры (2100-1750 до н.э.), в Тобольской и Ишимской долинах, к востоку от Уральских гор. Они были скотоводы - кочевники и связаны с Абашевской культурой (2500-1900 до н.э.), начиная с верхнего Дона, Волги до Урала, и культура Полтавка (2700-2100 до н.э.), простирается от Нижнего Дона, Волги до Каспийской низменности. Синташтинско-Петровкая культура была первой в Бронзовый век индо-европейцев к западу от Урала, открыв путь на обширные равнины и пустыни Центральной Азии, богатые металлами гор Алтая. Арийцы быстро распространились по всей Центральной Азии, от берегов Каспийского моря до Южной Сибири и Тянь-Шаня, через торговлю, сезонные миграция стад, грабежи и набеги.Военные колесницы для лошадей, вероятно, были изобретены людьми синташтинской культуры около 2100 до н.э. и быстро распространились на горной области Бактрии, Маргианы (современная граница Туркменистана, Узбекистана, Таджикистана и Афганистана). Медные рудники были интенсивно освоены на Урале, и прото-индо-иранцы Синташтинск-Петровской культуры в больших количествах экспортировали медб на Ближний Восток. Они, кажется, были привлечены к природным ресурсам Зеравшанской долины на Петровке, медно-добывающие колонии были созданы в около 1900 до н.э. Олово было особенно ценным ресурсом в конце бронзового века, когда оружие из сплава меди и олова, давало явное преимущество в военных действиях. В 1700-ых до н.э, индо-иранцы расширились до нижнего течения Амударьи в долину и поселились в сельскохозяйственных общинах. К 1600 году до н.э., старые укрепленные города Маргианы, Бактрии были заброшены, затоплены северными мигранты из степи. Группа культур Центральной Азии в индо-иранские времена известна как андроновский горизонт, и продолжалась до 800 г. до н.э.
Индоиранские миграции продвинулись дальше на юг через Гиндукуш. К 1700 г. до н.э., верховая езда скотоводов проникла в Белуджистан (юго-запад Пакистана). Долину Инда уступили около 1500 до н.э., в северной и центральной части Индийского субконтинента были переданы 500 лет до нашей эры. Западные миграции привели Старый индийский санскрит и езду боевых колесниц в Ассирию, где они стали правителями Митанни около 1500 до нашей эры. Мидии, парфяне и персы, все иранские докладчики из андроновской культуры, переехали на иранское плато около 800 г. до нашей эры. Те, кто остался в Центральной Азии помнят историю, как скифы, и потомки Ямной культуры, которые остались в Понтийско-Каспийской степи и стали известны как сарматы у древних греков и римлян.
Индоиранские миграции привели к высокой частоте R1a1-Z93 на юге Средней Азии, Иране и Индии. Наиболее высокая частота R1a1-Z93 (около 65%) достигается в кластере вокруг Кыргызстана, Таджикистана и севера Афганистана. В Индии и Пакистане, R1a колеблется от 15 до 50% населения, в зависимости от региона, этнической группы и касты. R1a, как правило, сильнее на северо-западе субконтинента, и слабее у дравидийцев Юг (Тамил-Наду, Керала, Карнатака, Андхра-Прадеш) и из Бенгалии на восток. В среднем около 40% брахманов (высшая каста в индуизме) принадлежат к R1a1-Z93, в некоторых местах и выше - вероятно в связи с эффектом основателя.
Материнские линии в Южной Азии, тем не менее, в подавляющем большинстве не-индо-европейские. Например, в Индии более 75% "родных" мт-ДНК M и R линий и 10%-Восточной Азии линий. В остаточных 15% гаплогруппы, примерно половина среднего происхождения Востока. Только около 7 или 8% может быть "северной" (Понтийско-Каспийских степей) происхождения, в основном в виде гаплогруппы U2 и W (хотя происхождение U2 по-прежнему обсуждается). Европейские линии мт-ДНК гораздо более распространены в Центральной Азии, и даже в Афганистане и севере Пакистана. Это говорит о том, что индоевропейские вторжения в Индию были проведены в основном мужчинами через войну, и первые крупные поселения их женщин были в северном Пакистане, Западной Индии (Пенджаб в Гуджарате) и на севере Индии (штат Уттар-Прадеш), где гаплогруппы U2 и W являются наиболее распространенными.
Тюркские линии и гаплогруппа R1a
Современные жители Центральной Азии, от Синьцзяна в Турции и от Волги до Гиндукуша, говорят в подавляющем большинстве на тюркских языках. Это может быть удивительно, но территориально это области, где индо-иранские ветви индо-европейской миграции расходились, от бронзового века андроновской культуры до железного века на скифской территории. Так почему же, что индо-европейские языки выживают только в славянской России, и в южной части Центральной Азии, в таких странах, как Таджикистан, Афганистан и некоторых частях Туркменистана? Почему же уйгуры, узбеки, казахи и кыргызы, у современных Понтийско-Каспийских степных народов (крымские татары, ногайцы, башкиры и чуваши) не говорят на индоевропейском языке? Генетически эти люди несут много R1a1-Z93, и в меньшей степени, линии N1b, C3, R1b1, N1c1, Q1. Это объясняется тем, что тюркские языки заменили иранские языки Центральной Азии предположительно между 4-м и 11-го веками нашей эры. Прото-тюркские возникли в Монголии и Южной Сибири вероятно у кочевых племен гуннов. Они принадлежит к алтайской языковой семье, как монгольский и маньчжурский (некоторые из них также включают корейский и японский, хотя они имеют очень мало общих слов в словаре). Пока неизвестно, когда прото-тюркские впервые появилась, но их распространение началось с гуннскими миграциями на запад через евразийские степи, и на всем пути в Европу, но остановилось на границах Римской империи.Гунны были потомками хунну. Тесты древней ДНК показали, что хунну были уже гибридны - Евразийского типа 2000 лет назад, со смешанными европейскими и Северо-Восточно азиатскими Y-ДНК и мт-ДНК. Современные жители провинции Xiongnu имеют примерно 90% монгольских линий против 10% европейских. Самое старое присутствие европейских мт-ДНК по Монголии и Байкала относится к более чем 6000 лет назад.
Похоже, что тюркский язык быстро заменил скифский и иранские диалекты всей Центральной Азии. Другие миграционные волны принесли больше тюркских генов в Восточной и Центральной Европе, как и хазары, авары, болгары и турки (=> см. 5000 лет миграций из евразийских степей в Европу). Многие из них были на самом деле из центрально-азиатских кочевников, которые приняли тюркский язык от своих восточных завоевателей, а также часть монгольских генов. Тюркские вторжения осуществлялись во многом с помощью R1a1-Z93.
Тюркские языки не сохранились в Европе за пределами Понтийско-Каспийских степей. Болгарский язык, несмотря на то, что назван в честь тюркского племени, на самом деле славянского происхождения с мягким тюркским влиянием. Венгерский, иногда ошибочно принимают за наследника гуннского из-за совпадения названия, на самом деле этот язык от уральских носителей N1c1 (мадьяров). Десятки тюркских языков в современном мире имеют высокую степень взаимопонимаемости в связи с их сравнительно недавним общим происхождением и кочевым характером миграций тюрков (до недавнего времени).
Греческие субклады гаплогруппы R1a1
Мало что известно о прибытии прото-греческого языка из степи. Микенская культура началось около 1650 г. до н.э. и, несомненно, является продолжением культуры степи. Тесная связь между микенскими и прото-индо-иранскими языками предполагает, что они разделились довольно поздно, в период между 2500 и 2000 годах до нашей эры. Археологически, микенские колесницы, наконечники копий, кинжалы и другие бронзовые предметы демонстрируют поразительное сходство с Сейсмино-Турбинской культурой (феноменом) (ок. 1900-1600 до н.э.) в северной русской лесостепи, известной большой подвижностью своих кочевых воинов (места находились так далеко, как Монголия). Поэтому вероятно, что микенцы спустились с территории современной России в Грецию в период между 1900 и 1650 до н.э., где они смешались с местным населением, чтобы создать новый уникальность греческой культуры.Гаплогруппа I
I - самая старая гаплогруппа в Европе и по всей вероятности, единственная, которая возникла там (кроме субкладов других гаплогрупп). Считается, что I прибыли из стран Ближнего Востока, как гаплогруппа IJ около 35000 лет назад, и превратились в гаплогруппу I примерно 25000 лет назад. Это означает, что кроманьонцы, скорее всего, принадлежали (как минимум частично) к IJ или I. В настоящее время гаплогруппа I составляет от 10 до 45% населения в большинстве стран Европы. Она разделена на четыре основных субклада.Самые ранние мегалитические сооружения (5000-1200 до н.э.) в Европе были построены мужчинами, принадлежащими к гаплогруппе I, затем к ним присоединились новички неолита, такие как G2a и E1b1b.
Гаплогруппа I1

Гаплогруппа I1 (ранее I1a) является наиболее распространенным субкладом I. Она локализована в основном в Фенноскандии, Северной Германии и Дании, где может представлять более чем 35% населения. Также в высокой концентрации у финнов и встречается от умеренной до низкой концентрации везде, куда вторгались германские племена и викинги.
В период неолита, предположительно I1 были частью успешной культуры Эртебеле (5300-3950 до н.э.) и Funnelbeaker культуры (4000-2700 до н.э.). Период шнуровой керамики (3200-1800 до н.э.) знаменует приход индоевропейских народов гаплогруппы R1a из украинских степей.
I1 определяется по крайней мере, 15 уникальными мутациями, которые указывают, что эта линия была выделена в течение длительного периода времени, или испытало серьезное "бутылочное горлышко". Хотя первое расщепление мутаций I1 от I2 могли возникнуть до 20 000 лет назад, люди, принадлежащие к этой гаплогруппе происходят от одного человека, который жил примерно 5000 лет назад. Это соответствует приходу индоевропейцев, предполагая, что высокий процент коренного населения I1 могли быть убиты новыми мигрантами.
Гаплогруппа I2

Гаплогруппа I2, возможно, возникла в центральной или юго-восточной Европе около 17000 лет назад и развилась в шести основных субкладах: I2a1a (M26), I2a1b (M423), I2a2a (M223), I2a2b, I2b (L416) и I2C (L596).
Субклад I2a1a (M26, L158, L159.1/S169.1) был известен как I1b2 до 2005 года, I1b1b в 2006-7 и I2a1 с 2008 по 2010 год. Распространен, главным образом на Сардинии и среди басков, и редко встречается за пределами Иберии, Западной Франции, на западном побережье Италии и Средиземноморского побережья Магриба. На его долю приходится около 40% всех Y-ДНК гаплогрупп среди Сардинии. Возраст I2a1a оценивается в 8000 лет.
I2a1b (M423, L178) был известен как I1b до 2007 года, и I2a2 с 2008 по 2010 год. Характерен на Балканах и в Карпатах, с максимальной частотой наблюдается среди Динарских славян (хорваты, сербы и боснийцы), а также в Молдавии и Румынии. Кроме того, в меньшей степени в Албании, Северной Греции, Болгарии, Словакии, Украине, Белоруссии и юго-западной России. Высокая концентрация I2a1b на северо-востоке Румынии, Молдовы и центральной Украине напоминает максимальное распространение Кукутень-трипольской культуры до ее поглощения индоевропейской культурой шнуровой керамики. Это может означать, что Кукутень-трипольская культура была родом из европейской группы охотников-собирателей, которые приняли сельское хозяйство после вступления в контакт с левантийскими фермерами, которые поселились на Балканах (гаплогруппы E-V13, J2b и Т) .
Современная территория I2a1a и I2a1b (Иллирия, Италия, Сардиния, средиземноморское побережье Франции и Испании) соответствует распространению неолитических культур (5000-1500 до н.э.), которые, как полагают, начался с приходом E -V13 и G2a фермеров и скотоводов из Фессалии (северная Греция). За ними последовали другие культуры (1500-1000 до н.э.) в бронзовом веке. R1b кельто-италики, как считается, пересекли Альпы и вторглись на итальянский полуостров около 1000 г. до н.э., заменив большинство коренных I2a, G2a и E-V13 людей (особенно в северной части полуострова).
Распределение гаплогруппы I2a1 (ранее I2a) в Европе
Субклад I2a2 (S33/M436/P214, P216/S30, P217/S23, P218/S32, L35/S150, L37/S153, L181) был известен как I1c до 2005 года и I2b до 2010 года. Предположительно, он связан кельто-германскими народами Северо-Западной Европы, теми, кто строил мегалиты (5000-1200 до н.э.). Разнообразие STR маркеров в I2b говорит о возрасте субклада около 13000 лет.I2a2 встречается во всех странах Западной Европы, и, видимо, они сохранились лучше после индоевропейских вторжений (=> см. выше R1b) в северной Германии, и восстановлены после германских вторжений в конце римского периода. В настоящее время I2a2 достигает пиков в центральной и северной Германии (10-20%), страны Бенилюкса (10-15%), а также на севере Швеции. Субклад также имеют от 3-х до 10% жителей Дании, Восточной Англии и северной Франции. Очень редко в Норвегии, что согласуется с тем, что она не была захвачена людьми из северной Германии.
Есть два основных субклада: I2a2a (M223) и I2a2b (L38/S154),которые в свою очередь подразделяются по крайней мере на 4 субклада каждый, хотя о них мало что известно. Субклад I2a2a1 (M284 +) встречается почти исключительно в Великобритании, где он предположительно и образовался около 3000 лет назад.
Распределение гаплогрупп I2a2 (ранее I2b) в Европе
I2b (L416, L417, L418) является очень незначительным субкладом и до сих пор наблюдается всего в нескольких образцах из Германии, Италии и Шотландии. I2C (L596, L597), вероятно, возник в районе Рейнской области. Его можно разделить на три группы: A, B и C. Группа A и B, предположительно, распространилась в кельтской экспансии бронзового и железного веков, наряду с R1b-S116/P312. Группа B географически ограничена в Германии, Швейцарии, Великобритании и Ирландии. Группа С имеет такое же распространение, но также была найдена во Франции, Италии и Норвегии. Группа B находится в низких частотах в Северо-Восточной Европе (Польша, Литва, Россия, Украина, Молдова), Юго-Восточной Европы (Румыния, Болгария, Крит) и Западной Азии (Анатолии, Грузии, Армении, Азербайджан и Северо-Западный Иран).Гаплогруппа N (Y-ДНК)
Гаплогруппа N распространена на Балтике, в Финляндии, на Урале и в Сибири, у балтов и славян, а также в умеренных частотах до Кореи и Японии. В Европе гаплогруппу N можно найти в очень высоких частотах среди современных финнов (68%), литовцев (45%), латышей (42%), эстонцев (38%) и у северных русских (35%). В умеренных частотах от 5 до 15% среди шведов, поляков, белорусов и украинцев.Гаплогруппа N, как полагают, возникла в Юго-Восточной Азии примерно 15000 до 20000 лет назад, но субклад N1c1 распространенный в Европе вероятно возник в Южной Сибири около 10 000 лет назад, и распространился в сторону Северо-Восточной Европы около 7000 лет назад. Вместе с миграциями отдельных субкладов гаплогруппы N1 распространялись нефритовые украшения в Евразии.
Гаплогруппа N1c1 связана с культурой Ямочно-Гребенчатой керамики (4200-2000 до н.э.), которая заложила основу в финских и прото-балтийских народах. Индоевропейская культура шнуровой керамики (3200-1800 до н.э.) постепенно захватила Балтийский регион и часть южной Финляндии около 2500 до нашей эры. Их слияние привело к гибридной культуре Kiukainen (2300-1500 до н.э.). Современные балтийские народы имеют примерно равные доли гаплогруппы N1c1 и R1a1 в результате древнего объединения уральских и прото-балтских культур.

Распределение гаплогруппы N1c1 в Европе
Продолжение
Комментариев нет:
Отправить комментарий